Questa piccola collezione di studi scientifici evidenzia come le cause che possono indurre le encefalopatie definite erroneamente come “autismo”hanno come denominatore comune un’origine di tipo neurotossico. Dal mercurio all’alluminio, ai vaccini coniugati, all’elevato livello di anticorpi per il morbillo nei vaccinati con MPR, alla presenza di DNA fetale, all’insorgenza di una o più condizioni autoimmuni a livello del sistema nervoso centrale, all’alterazione profonda dei rapporti di equilibrio del microbiota gastroenterico, il loro ruolo nel determinare disturbi neurologici di diversa natura e a vario livello di gravità è oramai accertato (Davide Suraci, alias autoimmunityreactions, alias yellowbrain).
Vaccini e autismo
Commentary–Controversies surrounding mercury in vaccines: autism denial as impediment to universal immunisation.
What is regressive autism and why does it occur? Is it the consequence of multi-systemic dysfunction affecting the elimination of heavy metals and the ability to regulate neural temperature?
A two-phase study evaluating the relationship between Thimerosal-containing vaccine administration and the risk for an autism spectrum disorder diagnosis in the United States
Abnormal measles-mumps-rubella antibodies and CNS autoimmunity in children with autism.
Uno studio pubblicato sul Journal of Biomedical Sciences ha stabilito che l’autoimmunità verso il sistema nervoso centrale può giocare un ruolo causale nell’autismo. I ricercatori hanno scoperto che poiché molti bambini autistici ospitano elevati livelli di anticorpi del morbillo, devono condurre uno studio sierologico sugli autoanticorpi anti-morbillo-parotite-rosolia (MMR) e della proteina basica della mielina (MBP). Hanno usato campioni di siero di 125 bambini autistici e 92 bambini controllati. La loro analisi ha mostrato un aumento significativo del livello di anticorpi MMR nei bambini autistici. Lo studio conclude che i bambini autistici hanno avuto una risposta anticorpale inappropriata o anormale alla MMR. Lo studio ha determinato che l’autismo potrebbe essere il risultato di un’infezione atipica del morbillo che produce sintomi neurologici in alcuni bambini. La fonte di questo virus potrebbe essere una variante del morbillo-virus o potrebbe essere il vaccino MPR.
La prima causa nota di autismo fu il virus della rosolia. Questa causa di autismo è nota dagli anni ’60. E il virus della rosolia è uno dei tre virus vivi nel vaccino MMR.
“… il virus della rosolia è una delle poche cause conosciute di autismo. ” Walter A. Orenstein, M.D. US as Assistant Surgeon General, Director National Immunization Program in a letter to the UK’s Chief Medical Officer 15 February 2002.
[*questo riferimento lo trovate nell’archivio web della pagina CDC – occorre cercare o scorrere la pagina per vedere il testo il testo originale in inglese: “rubella virus is one of the few known causes of autism“. Poiché i documenti citati nella pagina originale dal CDC come prove per l’assenza di collegamenti con il vaccino sono stati costantemente screditati, sembra che il CDC abbia deciso di rimuovere la pagina e sembra che qualcuno abbia eliminato anche le versioni archiviate della pagina dall’archivio web ].
“…rubella virus is one of the few known causes of autism…”
Robert E. Weibel, Vito Caserta, David E. Benor, Geoffrey Evans
viene rilevato:
Un totale di 48 bambini, dai 10 ai 49 mesi, hanno soddisfatto i criteri di inclusione dopo aver ricevuto il vaccino contro il morbillo, da solo o in combinazione. Otto bambini sono morti e il resto ha avuto regressione e ritardo mentale, convulsioni croniche, deficit motori e sensoriali e disturbi del movimento. L’insorgenza di segni o sintomi neurologici si è verificata con una distribuzione non casuale e statisticamente significativa dei casi nei giorni 8 e 9. Nessun caso è stato identificato dopo la somministrazione di parotite monovalente o vaccino contro la rosolia.
E conclude:
Questo raggruppamento suggerisce che una relazione causale tra il vaccino contro il morbillo e l’encefalopatia può esistere come una rara complicanza dell’immunizzazione del morbillo.
L’attenzione di numerosi ricercatori è sempre più orientata alla ricerca delle cause, dei fisiologismi e delle conseguenze dello stress ossidativo. Comprendere i fisiologismi del danno ossidativo significa modificare radicalmente sia le modalità di approccio diagnostico che quelle terapeutiche. In questa disamina viene evidenziata la natura dello stress ossidativo e le sue varie tipologie, focalizzando l’attenzione sulla serie limitata del DNA mitocondriale e sul fatto che i danni individuati in questa serie sono numericamente superiori a quelli individuati nel DNA nucleare. L’espressività genica viene fortemente condizionata da moltissimi fattori stressogeni e, tra questi, i componenti vaccinali. Quali effetti sul metabolismo cellulare? Quali conseguenze sul metabolismo dei tessuti, degli organi, dei sistemi? Quali implicazioni con il numero sempre crescente di patologie autoimmuni? A voi il percorso di conoscenza. (Premessa a cura di Davide Suraci, alias autoimmunityreactions, alias yellowbrain)
Tipi di danni ossidativi a mitocondri e mtDNA
[1]È possibile che si verifichino diversi tipi di danni ossidativi, di cui solo una [1a]serie limitata è stata specificamente individuata nel DNA mitocondriale, mentre si sospetta che la [1b]maggior parte di questo tipo di danni si verifichi nei mitocondri a causa delle condizioni ossidative della matrice. [1c]È noto da oltre un decennio che il livello stazionario del danno ossidativo nell‘mtDNA è superiore a quello dell‘nDNA: [1b]il DNA mitocondriale subisce 3-10 volte più danni del DNA nucleare a seguito di stress ossidativo. I danni endogeni ed ambientali influiscono quindi sul metabolismo del DNA mitocondriale che, avendo delle limitatissime capacità di autoriparazione, è maggiormente soggetto a mutazioni.
DNA Mitocondriale (mtDNA)
Nel
medesimo periodo di tempo, i ricercatori hanno identificato e
misurato varie forme di [2]danno
endogeno ed ambientale del DNA mitocondriale o mtDNA
–ossia
il DNA collocato all’interno dei mitocondri con 16.569 paia di basi
e 37 geni che codificano 13 polipeptidi, 22 tRNA e 2rRNA, che è
coinvolto nella produzione di proteine necessarie alla respirazione
cellulare–
esplicando i suoi percorsi di riparazione. È interessante notare che
i mitocondri [2a]sembrano
contenere solo una parte dei molteplici meccanismi di riparazione
dell‘nDNA,
o DNA
nucleare,
anche se il loro tipo di danno [2b]caratterizza
anche il DNA mitocondriale, la cui ridotta capacità di riparazione
può quindi, in parte, spiegare l’alta frequenza di mutazioni
del cromosoma mitocondriale
[1b]Negli
ultimi 15 anni è stato effettivamente evinto che nei mitocondri
mancano gli enzimi necessari per la riparazione dell’escissione
nucleotidica, la via principale per l‘eliminazione degli addotti al
DNA –in
genetica molecolare, un addotto al DNA è un frammento
del DNA covalentemente legato ad una sostanza chimica,
che potrebbe rappresentare l‘inizio
della formazione di cellule cancerose–
risultanti dall’esposizione ai raggi UV,
dall‘esposizione/ingestione a/di idrocarburi policiclici aromatici
cancerogeni ed agenti chemioterapici
[2c]A
differenza del DNA
nucleare, caratterizzato
da meccanismi multipli per la propria riparazione, i mitocondri
appaiono dunque limitati nella loro capacità di correggere
tutte le possibili forme di danno sia proprie che al DNA
mitocondriale
[3]Poiché
la replicazione dell’mtDNA
dipende dalla trascrizione genica, un suo eventuale danno può
alterare l’espressione
genica mitocondriale nei seguenti modi:
•
causa
[3a]errori
di incorporazione del neuclotide del DNA-polimerasi di tipo γ che
portano a mutazioni
Le
mutazioni possono essere spontanee, ereditarie da parte di madre,
oppure il risultato appunto di difetti del DNA nucleare presenti in
quei geni che mantengono il DNA mitocondriale
•
[3b]interferisce
con il priming della replicazione dell‘mtDNA per quel che concerne
RNA-polimerasi;
La
replicazione del DNA mitocondriale e la correlata trascrizione sono
strettamente correlate, in quanto il sistema per la
trascrizione mitocondriale fornisce le trascrizioni per i primer RNA
per la replicazione di DNA mitocondriale ed espressione genica
• induce [3c]mutagenesi trascrizionale oppure terminazione prematura della trascrizione
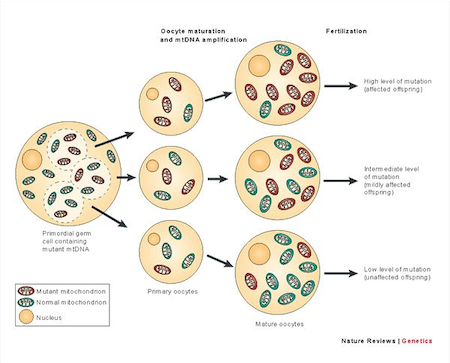
DNA mitocondriale mutato e DNA mitocondriale normale e come possono essere trasmessi alla discendenza tramite l’oocita con amplificazione del mtDNA.
L‘espressione genica mitocondriale è fondamentale per mantenere l’omeostasi cellulare: il suo controllo è unico in quanto i suoi componenti hanno doppia origine nei mitocondri (tutti gli RNA) e nel nucleo (tutti i fattori proteici). La regolazione della sintesi e della degradazione degli mtRNA, ossia RNA mitocondriali, determina i livelli stazionari delle proteine codificate a livello mitocondriale, permettendo così un accurato controllo del metabolismo energetico mitocondriale, creando quindi quelle basi affinché le cellule possano adattarsi alle variazioni degli stress ambientali e soddisfare le nuove richieste di energia cellulare. I difetti nell’espressione genica mitocondriale possono portare a disfunzioni della catena respiratoria con conseguente fenotipo di malattia multisistemica, che colpisce prevalentemente i tessuti muscolari e neuronali
[4]La replicazione e la funzione mitocondriale richiedono proteine codificate nel DNA nucleare; pertanto, la loro funzione nel complesso è sotto doppio controllo genetico
Ruolo e funzioni dei mitocondri
I [4]mitocondri ospitano il piccolo genoma circolare nelle cellule umane che è essenziale per la vita
Le
molteplici funzioni svolte dai mitocondri,
in particolare il loro [4a]coinvolgimento
nella regolazione della risposta immunitaria innata, ne fanno un
bersaglio privilegiato per i patogeni batterici in quanto
rappresentano una [4b]componente
chiave del metabolismo centrale attraverso la produzione e
la sintesi di adenosina trifosfato (in inglese Adenosine
Triphosphate o ATP) e di varie biomolecole
[4c]Tutte
le vie metaboliche della degradazione
ossidativa di
carboidrati, acidi grassi ed amminoacidi convergono in una tappa
finale, in cui l’energia prodotta dalle ossidazioni viene
utilizzata per la sintesi di adenosina
trifosfato o ATP,
che
si verifica in seguito al trasferimento degli elettroni, sottratti
durante le ossidazioni, all’ossigeno
Oltre
ad avere un ruolo cardine nella [4a]risposta
immunitaria innata, i mitocondri
partecipano anche alla [4d]segnalazione
cellulare attraverso la produzione di specie reattive
dell’ossigeno (in inglese reactive oxygen species o ROS)
che danneggiano i lipidi di membrana e le proteine, oltre al DNA
mitocondriale o mtDNA
ed infine il [4e]controllo
dell’omeostasi del calcio e il controllo della sopravvivenza
cellulare / morte cellulare programmata
Caratteristica
dei mitocondri
è che [5]l’ambiente
della matrice mitocondriale è piuttosto inospitale per le
macromolecole che vi risiedono. Sia la [5a]capacità
metabolica che la composizione della membrana dei mitocondri rendono
la matrice di questi organelli un probabile sito per la generazione
di una serie di aldeidi reattive. Le aldeidi, prodotte
fisiologicamente nei nostri tessuti, vengono secrete in aumentato
numero, ad esempio durante il metabolismo degli alcolici, e possono
entrare nell’organismo umano anche dall’ambiente, dove sono
presenti con altri agenti reattivi
Durante
la normale risposta immunitaria, [5b]i
mitocondri divengono promotori dell’infiammazione,
che svolge un ruolo cardine: [5c]l’infiammazione
è il tentativo dell’organismo di proteggersi da situazioni nelle
quali, senza essa ed i suoi effetti secondari che svolgono un ruolo
di limite, avverrebbero conseguenze più dannose
Il
suddetto concetto viene megnificamente esplicitato dal Prof.
Dr. George Vithoulkas insieme allo studioso ricercatore Stefano
Carlino nella loro pubblicazione scientifica dal titolo [5c]«The
“continuum” of a unified theory of disease»,
che porta in evidenza come l’organismo umano invii tutta una serie
di segnali (i c.d. sintomi)
in modo tale che essi possano essere accompagnati dolcemente,
piuttosto che soppressi, rispettando quindi l’organismo nella sua
interezza e totalità. Qualora si “sopprima” un sintomo,
evitando d’ascoltare il messaggio che esso porta, ecco che nel
tempo la serie di segnali aumenta in termini d’intensità e
profondità sino a coinvolgere in crescendo gli organi vitali più
importanti: in caso anche dopo tale fase questi segnali continuino ad
esser ignorati sopprimendone i sintomi, diverranno ulteriormente
intensi e prenderanno il sopravvento su tutto l’organismo
[5d]Lo
stress
ossidativo
–noto
come causa di induzione di allergie, patologie autoimmuni o
neurodegenerative con alterazione della crescita cellulare ed
infezioni croniche–
prodotto da un’infiammazione irrisolta e persistente (low
grade inflammation)
può esser un fattore importante coinvolto nel cambiamento della
dinamica delle risposte immunitarie, apportando alterazioni che
creano un caos immunologico con conseguente perdita di integrità
architettonica delle cellule e dei tessuti, portando a condizioni
croniche e/o neoplasia, cancro metastatico ed angiogenesi
Ad
ulteriore conferma del suddetto concetto vi è quanto scoperto nello
[5e]studio
scientifico «Stimulated
Human Mast Cells Secrete Mitochondrial Components That Have Autocrine
and Paracrine Inflammatory Actions»:
la somministrazione intraperitoneale di mtDNA isolato da cellule LAD2
nei ratti è stato rilevato nel loro siero entro 4 ore, indicando che
mtDNA extravascolare potrebbe entrare nella circolazione sistemica.
La secrezione dei componenti mitocondriali da parte dei mastociti
vivi stimolati può agire come “autopatogeno” contribuendo alla
patogenesi delle malattie infiammatorie
Le
molteplici sostanze chimiche presenti nell’ambiente, i metaboliti
dei componenti dietetici, i farmaci nelle terapie cliniche e/o in
generale le procedure mediche, oltre all’esposizione ai raggi UV
–si
noti che [Ib,
II]l’irradiazione
con la luce solare è
un fattore differente ed ha un ruolo importantissimo per la reazione
enzimatica e la [I]fosforilazione
ossidativa,
tematica qui di seguito approfondita–
sono fonti esterne di danni al DNA mitocondriale
[IIIa]I succitati elementi amplificano nell’organismo la produzione di lesioni endogene del DNA e generano addotti unici al DNA, oltre a – fra i molteplici possibili effetti consequenziali – inibire la trascrizione mitocondriale. Pertanto, nella valutazione dell’esposizione ambientale o della tossicità di una profilasi farmacologica, si dovrebbe considerare come possibile conseguenza la disfunzione dell’espressione genica mitocondriale o la mutagenesi dell’mtDNA
Danni al DNA nucleare versus DNA mitocondriale
[6]Mentre
il DNA nucleare è racchiuso in nucleosomi,
le molecole di mtDNA sono strettamente associate alla matrice
mitocondriale e formano strutture compatte quali i nucleoidi
A
differenza dei geni nucleari, ognuno dei quali ha spesso più
promotori dedicati,
[6a]tutti
i geni mitocondriali sono espressi insieme da tre soli promotori, che
sono riconosciuti dal meccanismo di trascrizione basale
mitocondriale: la polimerasi dell’RNA mitocondriale, e i fattori di
trascrizione mitocondriale Tfam e Tfb2m
I
nucleotidi
sono composti da complessi proteici che includono proteine coinvolte
nella replicazione e nella trascrizione, come i seguenti tre
promotori dei geni mitocondriali:
•
fattore
di trascrizione mitocondriale A, che svolge un ruolo
significativo nella replicazione dell’mtDNA,
nella trascrizione e nella struttura/organizzazione del nucleoide
mitocondriale;
•
polimerasi
gamma, che è coinvolto nel mantenimento della stabilità
genomica mitocondriale;
•
proteina
legante a singolo filamento mitocondriale (in inglese
single-strand binding protein o RPA), che
insieme al fattore di trascrizione
mitocondriale A ha il compito di sintetizzare una replica
esatta del genoma mitocondriale durante il processo di replicazione
Per
combattere i danni al DNA
nucleare,
i mitocondri possiedono percorsi di riparazione che comprendono
proteine
di riparazione codificate da geni nucleari che vengono trasportate ai
mitocondri.
È ancora piuttosto poco chiaro se questi enzimi di riparazione
preesistano nei mitocondri al momento del danno o vi si trasferiscano
in risposta ai segnali di danno
Le
vie di riparazione del DNA
nucleare
sono le seguenti:
•
[2a]inversione
diretta (in inglese direct
reversal
o DR)
tramite il controllo dell’attività
del gene glycosylasea uracile DNA (UNG) negli estratti mitocondriali;
•
riparazione
del disequilibrio (in inglese mismatch
repair
o MMR)
tramite la correzione dei disallineamenti nel DNA della polimerasi
-sintetizzato;
•
riparazione
della rottura del doppio filamento (in inglese double-strand
break repair
o DSBR)
tramite la replicazione dei filamenti di DNA principali e dei
filamenti di DNA in fase di sviluppo;
•
riparazione
dell’escissione di base (in inglese base
excision repair
o BER),
che inizia con riconoscimento
ed eliminazione di una base danneggiata oppure inadeguata da parte
della glicosilasi del DNA che scinde il legame N-glicosilico fra la
base ed il glucosio;
•
sintesi
della translesione (in inglese translesion
synthesis
o TLS),
comporta
il passaggio temporaneo dalle polimerasi δ e ε replicative alle
polimerasi a rischio di errore, come la pol η, con siti attivi
abbastanza grandi che possono ospitare lesioni del DNA, permettendo
così il loro bypass
Un
ampio [6b]spettro
di danni sia al DNA
nucleare
che al DNA
mitocondriale
viene arrecato dagli ossidanti liberi sia radicali che non radicali
generati all’interno delle cellule
In
tale range sono comprese [6c]piccole
modifiche alle basi, come 8-oxo-2′-deossiguanosina (8-oxo-dG) o
8-oxo-7,8-diidro-2′-deossiguanosina (8-oxodG) –sia
nel DNA
nucleare
che mitocondriale
è una delle forme predominanti di lesioni
ossidative
indotte dai radicali liberi ed è stata quindi ampiamente utilizzata
come biomarcatore per lo stress ossidativo e la carcinogenesi–
ed [6d]insulti
alla spina
dorsale zucchero-fosfato –importante
componente strutturale del DNA
nucleare
che si compone di 5 zuccheri carboni disossiribososio e gruppi
fosfato, estremamente importanti nella funzione del DNA
nucleare
stesso–
come siti
abasici,
anelli
ossidati di desossiribosio
e rotture
del filamento del DNA,
solo per citarne alcuni
[1b,
2c]Fino
ad oggi, solo una serie limitata di questi danni caratteristici sia
del DNA
nucleare
che del DNA
mitocondriale
sono stati specificamente rilevati in
quest‘ultimo,
ma si sospetta che la maggior parte di essi si verifichi nei
mitocondri,
a causa delle condizioni ossidative della matrice
mitocondriale
I
casi di danno dei [7]siti
abasici, dovuti principalmente alla perdita di purina dal DNA, si
verificano ad un livello stazionario di >50.000 siti per cellula
nei tessuti dei mammiferi. Il [7a]cervello,
ricco di mitocondri, ha il maggior numero di siti abasici
Il
2′-deossiriboso del sito abasico può essere [7b]ossidato
per formare zuccheri modificati e rotture di singoli filamenti. I
siti abasici e le rotture del filamento a filo singolo (in inglese
single
strand breaks
o SSBs)
sembrano essere le forme predominanti di danno all‘mtDNA
prodotto da H2O2
–perossido
di idrogeno,
noto anche come acqua
ossigenata,
è il più semplice dei perossidi–
e dallo stress ossidativo indotto dai rotenoni nelle cellule in
coltura
Oltre a danneggiare direttamente il cromosoma mitocondriale, [8]gli ossidanti biologici possono reagire con i nucleotidi liberi producendo substrati aberranti per la sintesi del DNA mitocondriale e dell’RNA: mentre i mitocondri possiedono un enzima in grado ad esempio di eliminare 8-oxo-2′-deossiguanosina (8-oxo-dG) dal gruppo di substrati, possono verificarsi e persistere anche altri danni ai nucleotidi
Fosforilazione e fotocatalisi
[I]La fosforilazione ossidativa –nelle cellule eucariotiche ha luogo nei mitocondri –corrisponde alla sintesi di ATP ed induce la generazione di [Ia]una serie di molecole reattive all’interno dei mitocondri, a causa della fuga di elettroni dalla catena di trasporto; [Ib]in tale contesto la fotocatalisi –sistema catalitico attualmente poco noto che applica l’irraggiamento solare e reazioni foto-chimiche– implica raggiungere/attivare nei mitocondri i processi di eccitazione degli atomi delle molecole di acqua presenti nei mitocondri stessi, decomponendone alcune molecole in ossigeno e idrogeno, rilasciando quindi elettroni
I processi metabolici nei mitocondri generano [3a]lesioni del DNA che è impossibile riparare ed interferiscono con la replicazione e la trascrizione dell’mtDNA per tutti quei tessuti altamente dipendenti dall’ossigeno e quindi dalla fosforilazione ossidativa, anche indirettamente correlati in quanto semplicemente beneficiano degli effetti conseguenti alla fosforilazione stessa, come [II]quelli di:
• cervello,
in specifico sistema nervoso centrale e periferico;
• muscolo cardiaco, scheletrico e muscolatura liscia in generis;
• pancreas, in specifico cellule β pancreatiche produttrici di insulina;
• reni
Al
contempo si evidenzia che [Ib,
III]negli
ultimi anni sono stati trovati numerosi enzimi che mostrano uno
spettro di reazione completamente diverso o esteso in conseguenza
alla fotocatalisi
, ossia all’irradiazione
con la luce solare,
in specifico a tipo, quantità e latitudine di irradiazione solare
giornaliera
Ad
oggi [IV]l’attivazione
fotochimica degli enzimi non è ancora diventata un approccio
standard: uno dei fattori limitanti è stata la cinetica
relativamente scarsa di trasferimento degli elettroni fotoeccitati
all’enzima, un altro è rappresentato dalla generazione di forti
ossidanti e radicali liberi reattivi
[8a]Una
degradazione dell’mtDNA
si verifica per compensare lo squilibrio del gruppo nucleotidico, al
fine di ripristinare i rapporti di nucleotidi necessari per una
corretta sintesi del DNA
mitocondriale stesso
I
genomi ed i loro nucleotidi precursori sono altamente esposti a
specie reattive dell’ossigeno (in inglese reactive oxygen
species o ROS), generati sia come sottoprodotti
della respirazione dell‘ossigeno o esecutori molecolari nella
difesa dell‘ospite, sia per esposizione ambientale a radiazioni
ionizzanti e sostanze chimiche
Il
[9]superossido,
non potendo diffondersi attraverso la membrana mitocondriale interna
(in inglese Inner Mitochondrial Membrane o IMM)
si limita alla matrice mitocondriale, dove può interagire con
le proteine Fe-S per liberar ferro / esser convertito in perossido di
idrogeno dalla dismutasi manganese-superossido, Mn-SOD
[9a]
Il perossido
di idrogeno, a differenza del superossido, può diffondersi
attraverso la membrana mitocondriale interna, eppure rimanendo nella
matrice può essere sottoposto alla chimica di Fenton con Fe(II) per
produrre radicali idrossili, altamente reattivi nei confronti del DNA
e degli acidi grassi polinsaturi (in inglese Polyunsaturated Fatty
Acids o PUFAs)
[9b]L’auto-ossidazione
dei legami insaturi negli acidi grassi polinsaturi porta alla
formazione di prodotti aldeidi distintivi che vengono regolarmente
neutralizzati dalla coniugazione del glutatione
o dall’attività dell’aldeide
deidrogenasi nei mitocondri
[10]Le
membrane esterna mitocondriale e quella plasmatica cellulare, i DNA
nucleare e mitocondriale possono essere esposti ad una diversa
miscela di aldeidi α,β-insaturi, ossidanti non radicali –i
più diffusi sono 4-HNE,
malondialdeide
(MDA),
acroleina
e crotonaldeide–
in grado d’indurre alterazioni delle proteine e del DNA
mitocondriale
nella matrice e di diffondersi verso l’esterno attraverso la
membrana
mitocondriale interna
Negli
ultimi anni sono stati studiati i [11]polimorfismi
mitocondriali del DNA e le mutazioni somatiche, al fine di
comprendere l’ereditarietà delle patologie umane, la longevità e
la migrazione delle popolazioni: alcune sequenze del cromosoma del
DNA mitocondriale sembrano essere instabili nel corso naturale
dell’invecchiamento, così come molte variazioni di singoli
nucleotidi nel mtDNA sono benigne, ma alcune favoriscono l’insorgenza
di patologie
La
[11a]neurodegenerazione
osservata durante sia l’invecchiamento che alcune patologie può
insorgere a causa di insulti multipli che impattano
sull’mtDNA, alcuni dei quali possono inibire la sintesi di pol γ
per indurre mutagenesi e delezioni di mtDNA
Le cellule [11b]possono contenere migliaia di copie del cromosoma mitocondriale, e prima che in una cellula sorgano fenotipi biochimici dev’essere compromesso un determinato livello di soglia dell‘mtDNA: parrebbe quindi esserci un [11c]controllo orchestrato delle condizioni della matrice mitocondriale, della dinamica mitocondriale e dei meccanismi di mantenimento dell‘mtDNA che permettono un cambiamento genetico naturale, prevenendo al contempo mutazioni e degradazione patogenetiche dell‘mtDNA
[1c]
Yakes and
Van Houten
–
Mitochondrial
DNA damage is more extensive and persists longer than nuclear DNA
damage in human cells following oxidative stress
|
doi:
10.1073/pnas.94.2.514
[3c]
D’Souza
and
Minczuk –
Mitochondrial
transcription and translation: overview
| doi:
10.1042/EBC20170102
[4]
Bhagavan
and
Chung-Eun Ha
– Essentials of Medical Biochemistry – The Human Organism: Organ
Systems, Cells, Organelles, and Our Microbiota | ISBN:
978-0-12-416687-5
[4a]
Walker
et
al.
– Powering the Immune System: Mitochondria in Immune Function and
Deficiency | doi:
10.1155/2014/164309
[4b]
Yaşar
Demirel
– Nonequilibrium Thermodynamics – Thermodynamics and Biological
Systems | ISBN:
978-0-444-59557-7
[4c]
Bhagavan
and
Chung-Eun Ha
– Essentials of Medical Biochemistry – Electron Transport Chain,
Oxidative Phosphorylation, and Other Oxygen-Consuming Systems
| ISBN:
978-0-12-416687-5
[4d]
Ray
et al.
– Reactive oxygen species (ROS) homeostasis and redox regulation in
cellular signaling | doi:
10.1016/j.cellsig.2012.01.008
[4e]
Marchi
et
al.
– Mitochondrial
and Endoplasmic Reticulum Calcium Homeostasis and Cell Death
| doi:
10.1016/j.ceca.2017.05.003
[5e]
Zhang
et
al.
– Stimulated
Human Mast Cells Secrete Mitochondrial Components That Have Autocrine
and Paracrine Inflammatory Actions
| doi:
10.1371/journal.pone.0049767
[6c]
Valavanidis
et
al.
– 8-hydroxy-2′
-deoxyguanosine (8-OHdG): A Critical Biomarker of Oxidative Stress
and Carcinogenesis
| doi:
10.1080/10590500902885684
Stimolazione di autoanticorpi post vaccinali indotta da vaccinazione in soggetti apparentemente sani.
Negli studi evidenziati negli screenshots, un riferimento scientifico sulla possibilità – accertata clinicamente – che tale induzione alla produzione autoanticorpale avvenga, oltre che in soggetti sani (primo screen), anche in soggetti portatori di patologie autoimmuni (secondo screen). Questi screen sono tratti da “Vaccines & Autoimmunity” del Dr Yehuda Shoenfeld et al. Wiley Editions, 2015. Nel primo screen si legge:
“….L’induzione di autoanticorpi è stata studiata in bambini sani successivamente a vaccinazione con vaccini antiepatite A e antiepatite B. L’induzione di autoanticorpi 6 anni dopo l’inoculazione del vaccino antiepatite B è stata studiata in 210 bambini immunizzati alla nascita con il vaccino antiepatite B ricombinante (Belloni et al., 2010)…”
Nel secondo screen una tabella in cui sono riportati i dati relativi agli autoanticorpi sviluppati in soggetti portatori di patologie autoimmuni a seguito di vaccinazioni antiinfluenzale, antipneumococcica e antipapilloma virus. Ragioni a pieno sostegno del rifiuto totale di qualsiasi vaccinazione.
Ulteriori conferme sui danni indotti dai vaccini mediante produzione di autoanticorpi:
“…Come negli esseri umani, l’induzione di autoanticorpi è possibile in seguito alla vaccinazione negli animali. Gli autoanticorpi sono più frequentemente presenti nei cani più vecchi che in quelli più giovani (Papini et al., 2005). Di conseguenza, quando i cani anziani vengono vaccinati e testati per gli autoanticorpi dopo la vaccinazione, è difficile definire quale sia una conseguenza diretta del vaccino e cosa si è accumulato durante la vita. Nel primo studio sperimentale controllato per studiare la produzione di autoanticorpi dopo la vaccinazione di routine, i ricercatori hanno arruolato giovani cani che non erano stati precedentemente vaccinati (Hogenesch et al., 1999).”
“…Hanno dimostrato che la vaccinazione obbligatoria contro la rabbia, il virus del cimurro canino e il parvovirus canino 9 ha provocato la produzione di svariati autoanticorpi…”
Autoanticorpi indotti dalle vaccinazioni nei cani
Già nel 2009 Toplak e Avcin avevano ipotizzato la produzione autoanticorpale indotta dai vaccini e la loro responsabilità nell’induzione di patologie autoimmuni.
Leggete che cosa concludevano:
“…La vaccinazione contro i microrganismi patogeni è uno dei principali risultati della medicina moderna, ma a causa di un numero crescente di segnalazioni di reazioni avverse la procedura di vaccinazione ha indotto anche un considerevole dibattito. È noto che alcune infezioni sono coinvolte nell’attivazione della produzione di autoanticorpi, che potrebbero portare a reazioni avverse autoimmuni in soggetti geneticamente predisposti.
Sulla base di questi risultati è stato ipotizzato che le vaccinazioni potrebbero indurre reazioni autoimmuni simili.
Allo stato attuale (ndr: era il 2009) non vi è alcuna prova evidente che le vaccinazioni siano associate a malattie autoimmuni conclamate, ma
è stato dimostrato che nelle persone geneticamente predisposte la vaccinazione potrebbe innescare la produzione di autoanticorpi e reazioni avverse autoimmuni.
I primi studi che osservavano la produzione di autoanticorpi dopo la vaccinazione sono stati condotti su cani e topi.
Diversi studi hanno successivamente dimostrato la produzione di autoanticorpi dopo la vaccinazione in pazienti con malattie autoimmuni,
ma ci sono solo dati limitati sulle risposte autoimmuni dopo le vaccinazioni in esseri umani apparentemente sani (ndr: il Dr Yehuda Shoenfeld confermerà nel 2015). Questa recensione riassume le prove attuali (ndr: anno 2009) sugli autoanticorpi indotti da vaccinazione in soggetti apparentemente sani, compresi gli studi su animali ed esseri umani.
Vuol dire reazioni e patologie autoimmuni presto e/o più avanti negli anni.. (Autoantibodies Induced by Vaccine – Nataša Toplak and Tadej Avcin – Department of Allergology, Rheumatology and Clinical Immunology, University Children’s Hospital, University Medical Centre Ljubljana, Ljubljana, Slovenia) – pp 93-102 of “Vaccines and Autoimmunity” by Yehuda Shoenfeld et al. Wiley Ed. 2015.
La similarità delle sequenze proteiche esistenti (e dimostrate)fra il proteoma umano e quello virale e batterico è notoriamente responsabile di numerose patologie/reazioni autoimmuni.
Due studi di Darja Kanduc citati nei seguenti miei articoli [Peptidi Batterici nel Proteoma Umano] e [Vaccini, Reazioni Autoimmuni e Barcode..] sulla co-evoluzione del proteoma umano e di quelli virali e batterici dovrebbero far riflettere attentamente tutti i medici, clinici e operatori sanitari che operano in “scienza e coscienza”.
“…I potenziali eventi avversi associati con le vaccinazioni per le malattie infettive sottolineano la necessità di analisi efficaci e la definizione dei possibili effetti collaterali derivanti dal vaccino. Utilizzando il proteoma HPV16 come modello, abbiamo quantificato i rischi effettivi e teorici della vaccinazione anti-HPV16, e definito lo spettro di malattia potenziale derivante da concomitanti reazioni crociate con le proteine dell’organismo umano…”
Citazione in (Journal of Experimental Therapeutics and Oncology – Vol. 8 – 2009, Vol. 8, pp. 65–76) – “Quantifying the possible cross-reactivity risk of an hPV16 vaccine”
“…Qualsiasi vaccino a base antigenica deve essere accuratamente progettato e criticamente selezionato al fine di evitare i potenziali effetti collaterali derivanti dalle similarità di sequenza a livello molecolare…”
“…Any antigen-based vaccine needs to be carefully and thoroughly designed and critically screened for potential side effects by comparing sequence similarity at the molecular level…”
“…Non esiste una chiara, definita, tabulazione matematica della possibile cross-reattività derivante dai rischi associati a un protocollo di vaccinazione…”
In altri termini, chi “progetta”(se si può usare questo termine)vaccini ignora totalmente l’esistenza della cross-reattività legata alla similarità delle sequenze proteiche che possono esserci fra le proteine del corpo umano e quelle antigeniche vaccinali. Il rischio esiste da sempre ma solo pochissimi hanno tenuto vivo questo filone di ricerca.
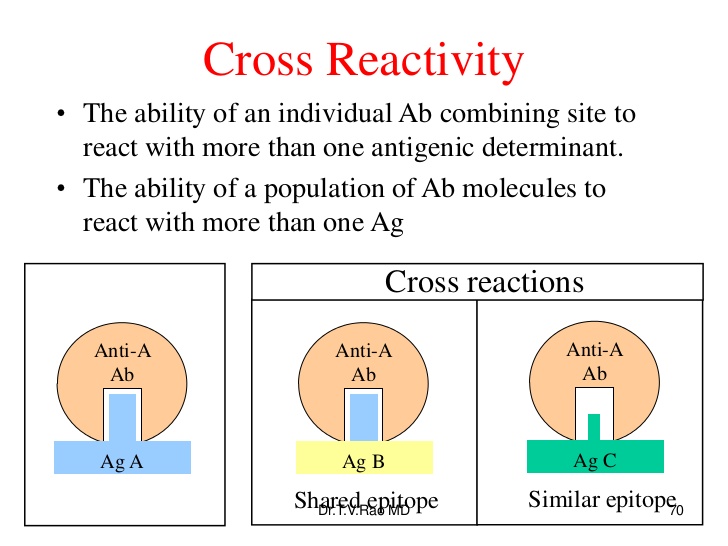
La cross-reattività fra gli epitopi e gli epitopi simili è causa di autoimmunità.
“Il potenziale carico patologico è pesante”... Con specifico riferimento all’antigene papilloma virus HPV16, “…le alterazioni delle proteine umane che ospitano le varianti HPV16 producono una serie di patologie che vanno dalle malattie muscolari ai disturbi neurologici. Di conseguenza, è logico postulare che il targeting di questi antigeni umani possa indurre molte delle sindromi riportate nella tabella allegata allo studio, scaricabile da qui: “Quantifying the possible cross-reactivity risk of an hPV16 vaccine” e di cui alleghiamo di seguito un estratto.
Citazione in: “Quantifying the possible cross-reactivity risk of an hPV16 vaccine” – (Journal of Experimental Therapeutics and Oncology – Vol. 8 – 2009, Vol. 8, pp. 65–76)
Dagli anni ’80 è stata introdotta una nuova generazione di vaccini singoli e obbligatori, comprendenti l’epatiteB, l’Hib, lo pneumococco, la varicella, il rotavirus e l’influenza, oltre a delle nuove strategie di marketing che meritano uno sguardo più attento.
Il vaccino per l’epatite B è un vaccino ricombinante, ciò significa che il suo DNA è stato geneticamente modificato rispetto a quello del virus da cui è derivato: insomma, un modo di dire dei produttori per sottolineare che non è più“vivo” e quindi non in grado di infettare alcuno. Finora, nessuno li ha contraddetti perché tecnicamente i virus sono “vivi” solo quando infettano le cellule di un ospite vivente.
Vaccino Epatite B Ricombinante.
Tuttavia, la “versione”OGM di questi virus potrebbe ancora possedere le caratteristiche di un virus, in quanto nessuno può dire con certezza quali malattie potrebbero essere originate da essa e, a maggior ragione, se queste possano assomigliare all’epatite B o meno.
Sostenere quindi che gli antigeni reingegnerizzati non siano “vivi” o “infettivi” è soltanto un’illusione. Il virus reingegnerizzato, inoltre, perde la sua piena antigenicità, il che obbliga all’aggiunta di sali di alluminio, formaldeide e proteine dei lieviti. (“Ingredients: Recombivax (Recombinant Hep B Vaccine, Merck)” 2011, vaccines.procon.org”)
I vaccini HPB – ancora oggi – rappresentano la premessa per gli studi di sicurezza per vaccini analoghi. Questi studi rinnegano la maggior parte delle reazioni/malattie autoimmuni(di fatto escludendole)per il semplice fatto che il momento in cui si manifestano spesso richiede settimane, mesi, anni.La“finestra temporale”entro cui esse si dovrebbero manifestare è pertanto ristretta(ore o qualche giorno)ed arbitrariamente determinata. Ciò ha portato le agenzie regolatrici a definire“sicuro”un vaccino che tale non è, nonostante esista un consistente corpus di articoli e di“case report”che lo associano a SIDS, autismo, sindrome di Guillain Barre, un’estesa gamma di danni cerebrali e neurologici – strettamente legati ai vaccini contenenti adiuvanti alluminio – oltreché ad un numero, vertiginosamente in crescita, di numerose patologie autoimmuni.
Una parte importante della morbilità dianzi citata è legata alla “politica” della somministrazione precoce. Nel 1991 il vaccino per l’epatite B venne reso obbligatorio, complice(e pretesto giustificatorio)il fatto che l’epatite B, C, l’AIDS ed altre malattie infettive si stavano diffondendo rapidamente attraverso le trasfusioni di sangue infetto.
La strategia adottata di vaccinare alla nascita per l’HPB“avrebbe dovuto”prevenire l’epatite B in tutti quei soggetti che, una volta diventati tossicodipendenti, avrebbero così avuto meno probabilità di contrarre il virus e avrebbero potuto così contribuire con le donazioni di sangue presumibilmente non infetto.
In non pochi congressi dedicati ai medici pediatri svoltisi in diverse parti del mondo emersero dubbi sulla necessità di somministrare precocissimamente un vaccino (quello per l’epatite B) i cui effetti avrebbero dovuto prevenire una malattia solo 25 – 30 anni dopo. Altri manifestarono(giustamente)il dubbio sull’eccessivo carico vaccinale nel primo anno di vita dei bambini (Pevsner, G., Letter, American Family Physician, January 1994, p. 47.). Non di meno, altri si chiedevano come mai esistessero delle contraddizioni sull’effettiva durata della“copertura”vaccinale(le documentazioni informative per i genitori su questo vaccino riportavano la copertura come a “vita”)e fossero necessarie – secondo altri articoli pubblicati – iniezioni successive di richiamo per mantenere alti i livelli di anticorpi. (Family Practice News, August 1, 1992, p. 23.)
Dopo circa cinque anni (inizio della seconda metà degli anni novanta) dall’avvento di questo vaccino – reso ormai obbligatorio – iniziarono a manifestarsi dei quadri clinici molto diversificati di patologie autoimmuni come mai si erano verificati in precedenza e la loro concomitanza con l’inizio dell’impiego nei vaccini di sali di alluminio, proteine antigeniche OGM non è da considerarsi una mera coincidenza.
Mi chiamo Dott.ssa Theresa Deisher. Sono fondatrice e scienziata capo del Sound Choice Pharmaceutical Institute, la cui missione è quella di educare il pubblico sulla sicurezza dei vaccini, nonché di fare pressione sui produttori affinché forniscano vaccini migliori e più sicuri per il pubblico. Ho conseguito il dottorato in Fisiologia molecolare e cellulare presso la Stanford University nel 1990 e ho completato il mio lavoro post-dottorato presso l’Università di Washington. La mia carriera si è svolta nel settore della biotecnologia commerciale e ho svolto il mio lavoro spaziando dalla scoperta di principi biologici di base e di farmaci allo sviluppo clinico.
Scrivo in merito a fatti scientifici inconfutabili sui contaminanti del DNA fetale nel vaccino contro morbillo-parotite-rosolia, che devono essere resi noti ai legislatori e all’opinione pubblica.
Il vaccino MPR II della Merck (così come il vaccino contro la varicella, il Pentacel e tutti i vaccini contenenti l’epatite A) è prodotto utilizzando linee cellulari fetali umane ed è pesantemente contaminato con DNA fetale umano dal processo di produzione. I livelli nei nostri bambini possono raggiungere fino a 5 ng/ml dopo la vaccinazione, a seconda dell’età, del peso e del volume sanguigno del bambino. È noto che quel livello attiva il recettore Toll-like 9 (TLR9), che possono causare attacchi autoimmuni.
Per illustrare la capacità autoimmune di piccolissime quantità di DNA fetale, considerate questo: il travaglio è innescato dal DNA fetale del bambino che si accumula nel flusso sanguigno della madre, innescando un massiccio rigetto immunitario del bambino. Questo è il travaglio.
Funziona così: frammenti di DNA fetalei di un bambino con circa 300 coppie di basi di lunghezza vengono trovati nel siero di una madre incinta. Quando raggiungono tra 0,46 e 5,08 ng/mL nel siero, innescano il travaglio tramite il meccanismo TLR9ii . I corrispondenti livelli ematici sono 0,22 ng/ml e 3,12 ng/ml. I livelli di DNA fetale in un bambino dopo essere stato iniettato con vaccini prodotti dal feto raggiungono lo stesso livello che innesca il rigetto autoimmune del bambino da parte della madre.
Chiunque affermi che il DNA fetale che contamina i nostri vaccini sia innocuo o non sa nulla di immunità e recettori Toll-like, oppure non sta dicendo la verità.
Se il DNA fetale può innescare il travaglio (una reazione autoimmune naturalmente desiderata), allora quegli stessi livellinei vaccini possono innescare l’autoimmunità in un bambino. Il DNA fetale frammentato contenuto nei vaccini è simile e (iii) dimensione, di ~215 coppie di basi.
Questa è una prova biologica diretta che i contaminanti del DNA fetale nei vaccini non sono in quantità basse e innocue. Sono un fattore scatenante proinfiammatorio molto forte.
La somministrazione di frammenti di DNA fetale umano (primitivo) non-self a un bambino potrebbe generare una risposta immunitaria che potrebbe anche reagire in modo incrociato con il DNA del bambino, poiché il DNA contaminante potrebbe presentare sezioni di sovrapposizione molto simili al DNA del bambino.
I bambini con disturbo autistico hanno anticorpi contro il DNA umano nella loro circolazione che i bambini non autistici non hanno. Questi anticorpi possono essere coinvolti negli attacchi autoimmuni nei bambini autistici. (iv)
La Duke University ha dimostrato in uno studio condotto di recente che sono stati osservati miglioramenti significativi nel comportamento quando i bambini con disturbo dello spettro autistico sono stati trattati con il loro sangue cordonale autologo conservatov . Questo trattamento mostra chiaramente che la maggior parte dei bambini con autismo non nasce con questo disturbo poiché malattie genetiche come la sindrome di Down o la fibrosi muscolare non possono essere curate con cellule staminali autologhe. Pertanto, un fattore scatenante ambientale, o fattori scatenanti, introdotti nel mondo intorno al 1980, quando l’autismo ha iniziato a crescere, devono essere identificati ed eliminati o ridotti nell’ambiente.
• Esiste una forte correlazione tra i crescenti tassi di autismo e il vaccino statunitense
passaggio produttivo dalle linee cellulari derivate da animali per il vaccino contro la rosolia alle linee cellulari umane abortite alla fine degli anni ’70.
• Il primo punto di cambiamento per l’anno di nascita del disturbo autistico (AD) è stato identificato nel 1981 per i dati della California e degli Stati Uniti, preceduto da un cambiamento nel processo di produzione:
o Nel gennaio 1979, la FDA ha approvato il passaggio di produzione per il virus della rosolia da un vaccino di origine animale (virus ad alto passaggio, HPV-77, coltivato ad esempio in cellule embrionali di anatra) alla linea cellulare fetale umana WI-38 utilizzando il ceppo virale RA27/3vii. Sia il vaccino monovalente contro la rosolia appena approvato sia un vaccino trivalente contro parotite, morbillo e rosolia utilizzano la linea cellulare fetale WI-38 per la produzione della porzione di vaccino contro la rosolia.
• Prima del 1980, il disturbo dello spettro autistico era una malattia molto rara, quasi sconosciuta. Secondo un aumento molto i dati del CDC, il tasso di autismo nel 2014 era di 1 su 59 bambini, dal momento che salì, rapidamente: nel 2000 era di 1 su 150. CDC: “I costi totali all’anno per i bambini con ASD negli Stati Uniti sono stati stimati tra $ 11,5 miliardi – $ 60,9 miliardi (dollari USA del 2011) (viii).”
• Di recente, sono state riconosciute duplicazioni e delezioni de novo fino al 10% dei disturbi dello spettro autistico simplex, corroborando i fattori scatenanti ambientali sulla genetica dei disturbi dello spettro autisticoix. • La porzione di rosolia del vaccino MPR contiene contaminanti del DNA fetale di origine umana pari a circa 175 ng, più di 10 volte la soglia raccomandata dall’OMS di 10 ng per dose di vaccino (x) .
• Nessun altro farmaco sul mercato riceverebbe l’approvazione della FDA senza un’accurata profilazione della tossicità (la FDA segue le linee guida internazionali ICH) -> questa non è mai stata condotta dall’industria farmaceutica per la contaminazione del DNA nel vaccino MPR.
• I vaccini prodotti con linee cellulari fetali umane contengono detriti cellulari e contaminanti DNA umano residuo, che non può essere completamente eliminato durante il processo di purificazione a valle del virus (xi). Inoltre, il DNA non è caratterizzato solo dalla sua sequenza (ATCG), ma anche dalla sua modifica epigenetica (ad esempio, modello di metilazione del DNA, ecc.). Questa decorazione è altamente specie-specifica, motivo per cui il DNA non umano verrà eliminato, mentre questo non è necessariamente il caso del DNA umano fetale.
Iniettare i nostri bambini con contaminanti di DNA fetale umano comporta il rischio di causare due patologie consolidate:
1) Mutagenesi inserzionale: il DNA umano fetale si incorpora nel DNA del bambino, causando mutazioni. La terapia genica basata sulla ricombinazione omologa di piccoli frammenti ha dimostrato che una quantità pari a 1,9 ng/ml di frammenti di DNA determina l’inserimento nel genoma delle cellule staminali nel 100% dei topi iniettati (xii). I livelli di frammenti di DNA fetale umano nei nostri bambini dopo la vaccinazione con vaccini contenenti morbillo, parotite, rosolia (MMR), Varivax (varicella) o epatite A raggiungono livelli superiori a 1,9 ng/ml. 2) Malattia autoimmune: il DNA umano fetale induce il sistema immunitario del bambino ad attaccare il suo stesso organismo.
Un’ulteriore preoccupazione: la contaminazione da retrovirus.
Il retrovirus endogeno umano K (HERVK) è un contaminante nel vaccino contro morbillo/parotite/rosolia (xiii).
• HERVK può essere riattivato negli esseri umani (xiv). Codifica una proteina (integrasi) specializzata nell’integrazione del DNA nel genoma umano.
• Diverse malattie autoimmuni sono state associate all’attività HERVK (xv). • Appartiene inoltre alla stessa famiglia di retrovirus del virus MMLV utilizzato in uno studio di terapia genica, in cui l’inserimento inappropriato del gene (mutagenesi inserzionale) ha portato a successive mutazioni somatiche aggiuntive e cancro in 4 ragazzi su 9 (xvi) .
• È quindi possibile che il frammento del gene HERVK presente nel vaccino MPR sia attivo, codifichi per l’integrasi o la proteina dell’involucro e abbia quindi il potenziale per indurre l’inserimento genico, favorendo la mutagenesi inserzionale e l’autoimmunità.
La presenza sia di DNA fetale contaminante ad alto livello sia di contaminazione da HERVK nel vaccino MPR rappresenta un rischio non studiato con enormi implicazioni e pericoli per la salute individuale e pubblica.
Soluzione: fare pressione sui produttori affinché tornino ai vaccini contro la rosolia derivati da linee cellulari animali, come era realizzato con successo in Giappone:
o Basato su ceppi Takahashi del virus della rosolia attenuato vivo, prodotto su cellule renali di coniglio. È stato recentemente dimostrato che una singola dose di questo vaccino mantiene l’immunità per almeno 10 anni quando la rosolia era sotto controllo regionale (xvii).
o Dividere il vaccino MPR in tre opzioni offerte individualmente, come avviene in Giappone.
È necessario modificare il processo di produzione del vaccino MPR per affrontare ed eliminare i rischi per la popolazione sopra menzionati.
Grazie per la vostra considerazione. Sarò felice di rispondere a qualsiasi domanda abbiate in merito a quanto sopra.
Sinceramente,
Dott.ssa Theresa A. Deisher
NOTE FINALI
i Lo et al. Am J Hum Genet. 1998 Apr;62(4):768-75 ii Enninga et al. Front Immunol. 2015 Aug 26;6:424 iii Deisher et al. Issues Law Med. 2015 Spring;30(1):47-70 iv Mostafa et al. 2014, J Neuroimmunol , Vol. 272, pp. 94–98; Mostafa et al. 2015, J Neuroimmunol , Vol. 280, pp. 16–20 v Dawson et al. Stem Cells Transl Med. 2017 May;6(5):1332-1339 vi Deisher et al. Issues Law Med, 2015 Vol. 30, pp. 25-46 vii https://www.cdc.gov/vaccines/pubs/pinkbook/rubella.html; Plotkin, SA. 2006, Clinical Infectious Diseases, Vol. 43, pp. S164–168; viii https://www.cdc.gov/ncbddd/autism/data.html ix Sebat et al. 2007, Science., Vol. 316, pp. 445-449; Sanders et al. 2011, Neuron, Vol. 70, pp. 863-885 x Series, WHO Technical Report. WHO EXPERT COMMITTEE ON BIOLOGICAL STANDARDIZATION 941; Deisher et al. Issues Law Med. 2015 Spring;30(1):47-70 xi Kramberger et al. Hum Vaccin Immunother. 2015;11(4):1010-21. xii McNeer, N A et al. “Systemic delivery of triplex-forming PNA and donor DNA by nanoparticles mediates site-specific genome editing of human hematopoietic cells in vivo.” Gene therapy vol. 20,6 (2012): 658-69. doi:10.1038/gt.2012.82 xiii Victoria et al. J Virol. 2010, Vol. 84, pp. 6033-6040 xiv Lee et al. PLoS Pathog. 2007 3(1):e10; Dewannieux et al. Biologicals, Vol. 38, pp. 366-70 xv Taietal.9,Nov2008, Mult Scler, Vol. 14, pp. 1175-80; Dickerson et al. 2008, Schizophr Res. 2008 Sep;104(1-3):121-6, Vol. 104, pp. 121-6 xvi Hacein-Bey-Abina et al. J Clin Invest. 2008 Sep;118(9):3132-42 xvii Jpn J Infect Dis. 2016 May 20;69(3):221-3
Attraverso l’iniezione nel torrente circolatorio di antigeni e di altri componenti vaccinali, si aggira la risposta immunitaria cellulo-mediata cioè quella, per intenderci, che si attiva quando un qualsiasi patogeno entra nel corpo umano attraverso le vie “canoniche” (mucose) in cui esiste un sistema immunitario “locale” perfettamente integrato con quello “sistemico”. Ciò non permette al sistema immunitario di produrre una risposta specifica (formazione di anticorpi) e mirata al vero patogeno. È l’inizio dell’innesco dell’autoimmunità.
Che cosa stiamo facendo? Stiamo letteralmente iniettando degli antigeni uniti ad adiuvanti negli esseri umani per stimolare “una” risposta immunitaria (quale?) e “una” produzione di anticorpi (quali?) in nome di quale strategia preventiva?
Dall’avvento delle vaccinazioni di massa è cresciuto il numero della persone che presentano costantemente una risposta immunitaria abnorme.
Sarebbe sufficiente un monitoraggio post vaccinale su larghissima scala per verificarlo. Eppure non interessa a nessuno.
L’inizio dell’autoimmunità
C’è molta differenza tra ciò che accade normalmente in un contesto di risposta immunitaria cellulo-mediata e ciò che accade quando si tenta di provocarla artificialmente nel tentativo di eludere la risposta medesima.
Si aggiunga inoltre che:
quando gli anticorpi intervengono per riconoscere e distruggere i tessuti/organi stessi del corpo a cui appartengono (autoimmunità) fagocitandoli, li portano inevitabilmente nel circolo sanguigno insieme ai loro residui di DNA. Ciò induce il sistema immunitario a produrre ulteriori anticorpi che vengono indirizzati a colpire ulteriormente i tessuti e/o gli organi oggetto dell’attacco primario in un ciclo reiterato.
Quindi, assistiamo ad un circolo vizioso didistruzione dei tessuti mediata da anticorpi, la cui conseguenza è un aumento del DNA cellulare nel sangue, innescando un aumento della produzione anticorpale , quindi una maggiore distruzione tissutale, e così via.
A causa di questo ciclo, le malattie autoimmuni non sono di solito auto-compensative. La via farmacologica tenta di arginare i sintomi, spesso fallendo, ma non interviene all’origine. Ecco “uno” dei perché i vaccini non servono a niente se non per indurre anche numerose patologie autoimmuni.
Il terreno(o suolo), inteso secondo il suo significato più ampio, è fondamentale nel determinare lo “stato di salute” di un ecosistema complesso quale quello costituito dalle molteplici e intricate relazioni fra substrato organico-minerale, forme di vita di vario ordine e natura, fattori abiotici (acqua, atmosfera, suolo).
In termini più “terra-terra”, un ecosistema complesso dotato di resilienza, omeòstasi e di biodiversità, è in grado di rispondere meglio alle sollecitazioni ambientali di varia natura ed entità.
Immaginate un prato naturale in cui vengano distrutti gli strati più superficiali di suolo costituiti da numerose e diverse specie di erbe e da una miriade di organismi e di microorganismi molto attivi nel mantenere un flusso in costante trasformazione di energia e di materia: piano piano, si aprono nel terreno delle ferite profonde che porteranno all’erosione e alla distruzione del microbiota (e del relativo microbioma) di quel terreno.
Verranno progressivamente alterate le strutture fisiche, chimiche, biologiche che avevano portato quel terreno ad evolversi fino all’innesco di processi di disfacimento strutturale e funzionale dell’originario ecosistema. Tutto il tratto gastro-intestinale è il nostro personale “terreno” che ci accompagna da tempo immemorabile ed è esattamente paragonabile all’ecosistema-prato naturale.
Se osservassimo al microscopio qualche grammo di terreno organico, lo vedremmo costellato da numerosissime forme di vita molto attive, esseri cellulari e pluricellulari al cui interno è possibile osservare un citoplasma che ricorda la consistenza della gelatina.
Analogamente, le cellule che costituiscono i tessuti del tratto gastroenterico – come i microorganismi che lo popolano – sono caratterizzate da un citoplasma dalla consistenza “gelatinosa”.
Quando una cellula è sana possiede questa caratteristica. I villi e i microvilli intestinali lo stesso. Se queste forme di vita, così come le nostre principali strutture di intercettazione e di assorbimento (villi e microvilli) a livello intestinale (e non solo) non potessero svolgere queste funzioni, si interromperebbero tutti quei processi di scambio nutrizionale e di detossificazione fondamentali per il mantenimento dell’omeòstasi.
Il gel intracellulare è costituito da proteine “strutturate” nell’acqua e capaci di trattenere e deviare le molecole tossiche eventualmente assorbite dai villi e dai microvilli.
Le cellule affiancate con villi e microvilli intestinali sani intercettano le tossine e le grandi molecole impedendone l’accesso diretto al flusso sanguigno.
Quando la struttura e l’integrità del citoplasma sono compromesse, le cellule si raggrinziscono, perdono la connessione reciproca parete-parete e si formano dei vuoti fra una cellula e l’altra. Attraverso queste lacune, riescono a transitare quelle macromolecole proteiche che non dovrebbero mai entrare nel flusso sanguigno.
Una volta nel sangue, il sistema immunitario deve poter neutralizzare queste macromolecole mediante la produzione di anticorpi. Questi anticorpi, spessissimo, interagiscono con i tessuti del corpo che li ha prodotti(cross-reattività) e sarà l’inizio di una malattia autoimmune.
In altre parole, il denominatore comune della malattia autoimmune può essere individuato proprio nell’intestino “che perde” (leaking gut) o permeabile (sindrome dell’intestino permeabile). E la causa dell’intestino permeabile è la perdita della turgidità delle sue cellule a causa della formazione di un gel tossico al loro interno, incapace di svolgere la sua primordiale funzione.
In questo studio di Darja Kanduc (*) pubblicato il 7 Novembre 2018 viene confermata l’esistenza di una co-evoluzione e di una correlazione evolutiva fra il proteoma umano e i proteomi virali, rispetto ad un suo precedente studio sul confronto peptidico umano e virale (Vedi: Peptidi Batterici e Virali nel Proteoma Umano…) (Kanduc et al., 2008c) e rimarcato ulteriormente che l’attuale prassi di progettazione dei vaccini ignora completamente l’esistenza delle reazioni crociate fra i proteomi virali e batterici e il proteoma umano e il mancato riconoscimento del “barcode” proteico da parte del sistema immunitario verso gli antigeni vaccinali implica numerosi dubbi sulle immunizzazioni artificiali a causa della possibilità di sviluppare reazioni autoimmuni e patologie su base autoimmune.
Isomerasi Umana – (By →AzaToth – based on 1wyi (http://www.pdb.org/pdb/explore/explore.do?structureId=1WYI), made in pymol, Public Domain, https://commons.wikimedia.org/w/index.php?curid=1007670)
I dati di somiglianza sequenziale e i saggi immunologici convergono nel descrivere “un sé umano“che si è evoluto dalle proteine virali e batteriche usando il cito-plasma arcaico come piattaforma metabolica. Oggi, al di là delle ovvie difficoltà di ricostruire un processo che affonda le sue radici in milioni di anni fa e che si sviluppò in condizioni chimico-fisiche lontane da quelle attuali, un tale contesto evolutivo indica che virus e cellule mammaliane probabilmente si sono evoluti da un comune pool di sequenze primordiali e in questo modo offre l’unica spiegazione logica per la massiccia condivisione di peptidi che collega le proteine virali e umane(Kanduc et al., 2008c).
Immunologicamente, un’interazione così vecchia di milioni di anni è protetta dall’immunotolleranza umana,così che il sistema immunitario agisce per difendersi dagli agenti infettivi mentre protegge l’integrità dell’ospite umano.
Infatti, sembra cheil sistema immunitario dei mammiferi riesca a evitare il rischio di crossreattività intrinseco alla condivisione di sequenze di peptidi con proteomi virali e microbici.
In realtà, un solido insieme di dati sperimentali(Kanduc et al., 2004, 2007, 2008a, b, Polimeno et al., 2008; Kanduc, 2009, 2010a, b, c, 2012b; Lucchese et al., 2009a, b, 2010, 2012a, b; Stufano et al., 2010; Novello et al., 2012)mostrano che, a seguito dell’infezione, le risposte immunitarie anti-patogeni sono generalmente dirette contro sequenze peptidiche epitopiche con scarsa o nessuna somiglianza con il proteoma dell’ospite .
Quindi, lo scenario evolutivo descritto non solo potrebbe migliorare la nostra comprensione dei fenomeni autoimmuni, ma getta anche un’ombra sulle attuali pratiche di immunizzazione e suggerisce che solo le immunizzazioni basate su piattaforme di peptidi che specificatamente indicano agenti infettivi con codice a barre colpiscono in modo specifico i patogeni senza il rischio di reazioni crociate autoimmuni nocive contro le proteine dell’ospite umano.
Docente di Biochimica II presso il Corso di Laurea Magistrale in Scienze Biosanitarie, Dipartimento di Bioscienze, Biotecnologie e Biofarmaceutica. Già docente del corso di Biochimica Comparata, presso il Corso di Laurea in Scienze Biologiche Vecchio Ordinamento. Componente del Dottorato di Ricerca in “Morfometria Analitica e Modelli di Medicina Molecolare” avente sede presso il Dipartimento di Anatomia Patologica della Facoltà di Medicina, Università di Bari. Già componente del Dottorato di Ricerca in “Cancerogenesi, Immunoregolazione ed Invecchiamento” avente sede presso il Dipartimento di Medicina Interna, Sezione di Geriatria, della Facoltà di Medicina, Università di Bari.
[1]Anergia, ovvero quando il sistema immunitario risponde con una “non-infiammazione” dei tessuti, ovvero uno stato di immunoresponsività, ossia di inattivazione o assenza d’infiammazione durante una normale risposta immunitaria. Perché il sistema immunitario in sviluppo dunque minimizza la produzione di citochine e chemochine pro-infiammatorie e la produzione di cellule linfocitarie di tipo T rispetto a quella delle cellule linfocitarie di tipo B? Ci sarà pure una ragione per cui il sistema immunitario in età pediatrica è in grado di modulare la propria risposta antiinfiammatoria – di fatto neutralizzandola – nei confronti del microbiota (soprattutto a livello intestinale)? Dunque un “fattore di tempestività” (il sistema immunitario infantile che si “concilia” con il microbiota intestinale lasciandogli fare il suo lavoro preparatorio – il terreno – al futuro sistema immunitario) pre-ordinato dalla genetica, unito ad un“fattore di opportunità” (una dichiarazione di pace e di cooperazione finalizzata ad una coesistenza reciproca) per il mantenimento, in definitiva, di un’omeòstasi duratura fra il corpo umano e i miliardi di microorganismi ospitati nel suo microbiota
Anergia, nota anche come anergia clonale
In presenza di stimoli microbici, i macrofagi intestinalipresenti nella mucosa intestinale [2] attuano una produzione ‘minimizzata’, ossia quasi pressoché nulla di citochine in contemporanea alla[3] sotto-regolazione di chemochine pro-infiammatorie: è interessante constatare come la mucosa dell’intestino tenue(o piccolo intestino)sia caratterizzata dall’[1]assenza d’infiammazione, nonostante la costante esposizione a batteri immunostimolanti ed a prodotti batterici in generis
Il sistema immunitario in sviluppo è quindi caratterizzato dalla produzione [4, 5] ‘minimizzata’ di citochine pro-infiammatorie e dallo sviluppo ‘asimmetrico’ delle cellule linfocitarie di tipo T rispetto a quelle di tipo B a favore di risposte regolatorie
Nonostante tale risposta immunitaria così ‘smussata’ potrebbe portare ad un’elevata suscettibilità alle infezioni, tale connubio regolatorioassicura che la colonizzazione dei microorganismi nel tratto gastro-intestinale avvenga senza rilascio di citochine/chemochine pro-infiammatorie, e quindi in[1]anergia, ossia con la caratteristica dell’inattivazione o assenza d’infiammazione
[6] La mucosa intestinale contribuisce quindi a mantenere l’omeostasi, al fine di prevenire un’auto-infiammazione tramite l’attivazione di una profonda sottoregolazione (o regolazione negativa) nel rilascio (avvenente appunto solamente dai 7 anni d’età in poi) di citochine e chemochine pro-infiammatorie da parte dei macrofagi
[2]Tale sottoregolazione in ambito intestinale si estende anche ai recettori Toll-like (in inglese Toll-like Receptor o TLR ) – indotti dal rilascio delle citochine stesse – in quanto tali recettori sono macrofagi intestinali appartenenti alla categoria dei recettori Toll-like, nello specifico TLR3-TLR9. Tale sottoregolazione, estesa ai recettori Toll-like, appartenenti ad una serie di profili molecolari microbo-associati, è caratterizzata a sua volta dall’[1]anergia, ossia dell’inattivazione o assenza d’infiammazione
La sottoregolazione da parte dei macrofagi intestinali per la produzione di interleuchine o IL[2a] interessa anche quella di liposaccaridi e persino quella di IgA, anch’essa atta a promuovere e mantenere un ambiente antinfiammatorio
[2, 7-8b] Molteplici studi scientifici hanno confermato come nell’organismo umano, sino ai 7 anni, sia innaturale la presenza di un elevato numero di citochine e chemochine, e come invero vi sia[9-11] interconnessione della regolazione immunitaria, che include la modulazione del sistema immunitario innato – compresi macrofagi, cellule dendritiche e l’induzione di cellule T regolatorie – ed è atta a mantenere l’omeostasi microbica e ad indurre meccanismi di difesa antimicrobica promuovendo un ambiente antinfiammatorio, e quindi a favore dall’[1]anergia
Ci si dovrebbe quindi domandare allora:
perché introdurre nell’organismo di un infante in crescita, direttamente nel sangue – bypassando quindi di fatto tutte le naturali barriere difensive – una qualsivoglia sostanza non bio-compatibile che oltretutto altera le naturali fasi dello sviluppo umano, come dovrebbe avvenire in maniera scientificamente dimostrata, ossia caratterizzata da anergia sino ai 7 anni d’età ? Ecco come viene annientato uno dei primi meccanismi difensivi/adattativi/conciliativi: l’anergia
[2] Smythies et al. Inflammation Anergy in Human Intestinal Macrophages Is Due to Smad-induced I B κ α
Expression and NF- B Inactivation κ | doi: 10.1074/jbc.M109.069955
[3]Andrew M. Platt & Allan McI. Mowat. Mucosal macrophages and the regulation of immune responses in the intestine | doi: 10.1016/j.imlet.2008.05.009
[4] PrabhuDas et al. Challenges in infant immunity: implications for responses to infection and vaccines | doi: 10.1038/ni0311-189
[7] Smythies et al. Human intestinal macrophages display profound inflammatory anergy despite avid phagocytic and bacteriocidal activity | doi: 10.1172/JCI200519229
[8] Smith et al. Intestinal macrophages lack CD14 and CD89 and consequently are down-regulated for LPS- and IgA-mediated activities. | doi: 10.4049/jimmunol.167.5.2651
[8a] Dupont et al. Antimicrobial peptides and the enteric mucus layer act in concert to protect the intestinal mucosa | doi: 10.1136/gutjnl-2014-307150
[8b] Million et al. New insights in gut microbiota and mucosal immunity of the small intestine | doi: 10.1016/j.humic.2018.01.004
[11] Elshar and Begun. The role of barrier function, autophagy, and cytokines in maintaining intestinal homeostasis | doi: 10.1016/j.semcdb.2016.08.018
Glossario
Cellule denditriche: sono cellule che appartengono al sistema immunitario con la funzione di presentare l’antigene ai linfociti B e T. Le cellule dendritiche nascono dalle cellule staminali emopoietiche nel midollo osseo e sono le più importanti dell’insieme delle APC (Antigen presenting cell), insieme ai macrofagi e ai linfociti B. Il loro nome deriva dalla particolare forma ramificata che assumono. Prima della maturazione sono presenti nei tessuti non linfoidi, e in particolare dove è più comune incontrare antigeni (per es., la cute e le mucose)
L’incontro con l’antigene dà inizio alla maturazione delle cellule dendritiche, legata anche a segnali chimici molto precisi, e in particolare alla presenza delle molecole di mediatori dell’infiammazione. Dopo l’incontro, l’antigene viene internalizzato per fagocitosi e processato da parte della cellula; quest’ultima migra dal tessuto non linfoide verso gli organi linfatici secondari (linfonodi, milza), le proteine antigeniche ottenute con la degradazione del patogeno vengono quindi esposte al fine di presentare l’antigene ai linfociti T
Cellule linfoidi o linfociti : sono cellule presenti nel sangue che costituiscono tra il 20 e il 40% dei leucociti (secondo i dati delle formule leucocitarie riconosciute). Hanno la dimensione di 7-15 micrometri (piccoli linfociti fino a 8 micrometri e grandi linfociti da 9 in su), con un nucleo rotondeggiante, un citoplasma scarso che si riduce a un sottile anello e pochi granuli, al microscopio elettronico è possibile distinguere un nucleolo (contrariamente a quanto si pensasse precedentemente). Svolgono un ruolo molto importante nel sistema immunitario, essendo la struttura portante della nostra risposta immunitaria adattativa (ovvero specifica per un tipo di antigene)
I linfociti derivano dalla linea linfoide delle lt, cellule staminali multipotenti presenti nel midollo osseo e a seconda del luogo, all’interno dell’organismo, nel quale avviene la maturazione cellulare, si ottengono due linee linfocitarie ben distinte: i linfociti B (da Bursa, con riferimento alla ‘borsa di Fabrizio’ scoperta da Girolamo Fabrici d’Acquapendente negli uccelli e dove si sviluppano i linfociti B ) e i linfociti T (dalla ghiandola del timo). Il luogo di maturazione dei linfociti risulta appunto diverso a seconda delle caratteristiche che questi hanno: il linfocita T infatti matura pienamente nel timo, mentre i linfociti B e NK (Natural Killer, i nostri linfociti “ancestrali”) hanno piena maturazione nel midollo osseo, ovvero dove nascono
Chemochine: sono particolari citochine (➔citochina) importanti nella dinamica di processi infiammatori, sono prodotte da varie cellule quali i macrofagi, i linfociti, i monociti e le piastrine ed esplicano la loro azione su recettori di membrana in grado di legare diverse chemochine fra loro (una stessa molecola di chemochina può combinarsi con più recettori). Sono le proteine responsabili del richiamo delle cellule dell’immunità –fra cui i monociti ed i linfociti– che mediano la risposta immunitaria innata e anche quella acquisita. Sono una sottoclasse di citochine
Citochine: sono molecole di natura proteica, sintetizzate dalle cellule dell’immunità: ad es., i macrofagi sintetizzano le citochine subito dopo il riconoscimento del profilo estraneo e alla fagocitosi. Le citochine hanno il ruolo di mediare la c.d. infiammazione, attraverso il richiamo di altri tipi cellulari, ed in questo caso si definiscono chemochine. Un altro compito delle citochine è di carattere sistemico poiché agiscono sull’ipotalamo, modulando l’innalzamanto della temperatura corporea
Controllano inoltre l’infiammazione, la risposta immunitaria, la crescita e il differenziamento cellulare in quanto mediatori intercellulari
Interleuchine: sono proteine di piccole dimensioni, appartenenti alla famiglia delle citochine (➔citochina) , che vengono secrete dai leucociti durante la risposta immunitaria. Promuovono la differenziazione e la proliferazione di determinate popolazioni di linfociti
Macrofagi: sono cellule del sangue e dei tessuti connettivi, di dimensioni relativamente grandi, dotati di proprietà fagocitarie, grazie alle quali, con meccanismi enzimatici, fagocitano, digeriscono ed eliminano protozoi, cellule e loro detriti, tessuti o sostanze estranee all’organismo. Svolgono un ruolo essenziale in immunologia poiché in grado di presentare l’antigene
Monociti: si formano nel midollo osseo, passano nel sangue e raggiungeno i tessuti dove matura a macrofago. I monociti macrofagi sono cellule importanti per la difesa immunitaria
Omeòstasi: è la tendenza naturale al mantenimento di un relativo stato di equilibrio interno delle proprietà chimico-fisiche di un organismo. I meccanismi omeostatici agiscono a livello delle cellule, dei tessuti e degli organi e riguardano diversi parametri: la temperatura corporea, il pH del sangue, il battito cardiaco, la pressione sanguigna, la concentrazione di glucosio nel sangue (o glicemia), e molti altri