Sostengono che le linfe insegnino al corpo come combattere un agente patogeno, ma questo studio mostra che i reni lo fanno. http://aac.asm.org/content/51/5/1700.full
Anche i muscoli fanno parte del sistema immunitario. Si noti che il danno da vaccino comporta quasi sempre problemi muscolari. http://dmm.biologists.org/content/9/6/697
I nostri batteri intestinali modulano il nostro sistema immunitario e allenano le nostre cellule B.
Traduzione e adattamento in lingua italiana a cura di Davide Suraci
Il Ruolo degli IFN di Tipo 1 (approfondimento)
Gli IFN di tipo I svolgono un ruolo essenziale nella lotta alle infezioni virali e le carenze nella segnalazione dell’IFN di tipo I sono state associate a scarsi risultati da COVID-19 in più studi. Questi casi sono spesso associati ad autoanticorpi contro gli IFN di tipo I. Come rivisto di seguito, gli IFN di tipo I sono stati utilizzati con un certo successo nel trattamento del COVID-19 grave, in particolare se somministrati molto presto nel processo patologico. Se, come sostenuto in precedenza, i vaccini a mRNA interferiscono con la segnalazione dell’IFN di tipo I, ciò potrebbe portare a una maggiore suscettibilità al COVID-19 nelle due settimane successive al primo vaccino, prima che sia iniziata una risposta anticorpale. Le cellule infettate da un virus rilevano la presenza della replicazione del virus attraverso una serie di recettori di riconoscimento del modello (PRR), che fungono da sentinelle che rilevano le strutture aberranti dell’RNA che spesso si formano durante la replicazione virale. Questi recettori rispondono oligomerizzando e successivamente inducendo IFN di tipo I, regolando in ultima analisi un gran numero di proteine coinvolte nella soppressione della proliferazione virale (Janeway e Medzhitov, 2002).
Uno studio multi-autore condotto da ricercatori a Parigi, in Francia, che ha coinvolto una coorte di 50 pazienti COVID-19 con vari gradi di gravità della malattia, ha rivelato che i pazienti con malattia grave erano caratterizzati da una risposta IFN di tipo I altamente compromessa(Hadjadj et al., 2020).Questi pazienti sostanzialmente non presentavano IFN-β e bassa produzione e attività di IFN-α.Ciò era associato a una carica virale ematica persistente e a una risposta infiammatoria esacerbata, caratterizzata da alti livelli di fattore di necrosi tumorale α (TNF-α) e Il-6. Gli autori hanno proposto la terapia con IFN di tipo I come potenziale opzione terapeutica. Un documento di diversi ricercatori negli Stati Uniti ha anche identificato una risposta infiammatoria unica e inappropriata in pazienti con COVID-19 grave, caratterizzata da bassi livelli di IFN sia di tipo I che di tipo III insieme a chemochine elevate ed elevata espressione di Il-6(Blanco- Melò et al., 2020). Gli IFN di tipo I sono stati persino proposti come opzione terapeutica per il COVID-19 grave. In un modello di criceto, i ricercatori hanno esposto i criceti alla SARS-CoV-2 e hanno indotto una risposta infiammatoria nei polmoni e un’infiammazione sistemica nei tessuti distali. Hanno scoperto che la somministrazione intranasale di IFN-α ricombinante ha comportato una riduzione della carica virale e un alleviamento dei sintomi (Hoagland et al., 2021). Uno studio di coorte retrospettivo su 446 pazienti affetti da COVID-19 ha determinato che la somministrazione precoce di IFN-α2b era associata a una ridotta mortalità intraospedaliera. Tuttavia, la terapia tardiva con IFN ha aumentato la mortalità e ritardato il recupero, rivelando che la somministrazione precoce della terapia con interferone è essenziale per una risposta favorevole (Wang et al., 2020a). Un numero sorprendente di persone ha autoanticorpi neutralizzanti contro gli IFN di tipo I, sebbene l’eziologia alla base di questo fenomeno non sia stata compresa. Uno studio che ha utilizzato la profilazione longitudinale di oltre 600.000 cellule mononucleate del sangue periferico e il sequenziamento del trascrittoma di 54 pazienti con COVID-19 e 26 controlli ha rilevato una notevole mancanza di risposte geniche stimolate dall’IFN di tipo I nelle cellule mieloidi di pazienti con malattia critica (van der Wijst et al. al., 2021).
Human Interferon – By Nevit Dilmen – Self created from PDB entry with Cn3D Data Source: https://www.ncbi.nlm.nih.gov/Structure/, CC BY-SA 3.0, https://commons.wikimedia.org/w/index.php?curid=1307713
Gli autoanticorpi neutralizzanti contro gli IFN di tipo I sono stati trovati nel 19% dei pazienti con malattia critica, nel 6% dei pazienti con malattia grave e nello 0% dei pazienti con malattia moderata. Un altro studio con sede a Madrid, in Spagna, ha rivelato che il 10% dei pazienti con grave malattia da COVID-19 aveva anticorpi autoimmuni contro gli IFN di tipo I (Troya et al., 2021). Uno studio multi-autore con sede in Francia ha rilevato che la mortalità per COVID-19 era significativamente più frequente nei pazienti con autoanticorpi neutralizzanti contro l’interferone di tipo I rispetto a quelli senza anticorpi neutralizzanti(55% vs. 23%)(Chauvineau ‐ Grenier et al., 2022) . Infine, Stertz e Hale(2021) notano che, a causa di autoanticorpi o forse di polimorfismi con perdita di funzione associati ai geni del sistema dell’interferone, le carenze nella produzione di interferone sono associate a ben il 15% di tutti i casi di COVID-19 potenzialmente letali .
mRNA Vaccinale non Prevede le Strategie di Metilazione?
Le strategie di metilazione per il mantenimento cellulare sono generalmente omesse dagli mRNA del vaccino? La metilazione degli mRNA è stata evolutivamente concepita per controllare la traduzione delle trascrizioni e quindi l’espressione dei geni da parte di una complessa cascata di proteine metilatrici (scrittori), de-metilatrici(cancellatori) e lettrici. La metilazione dell’adenosina è la modifica epitranscriptomica più abbondante dell’mRNA e si verifica in più siti attraverso la molecola dell’mRNA(Zaccara et al., 2019).
Una metilazione chiave dell’adenosina “N6-metiladenosina (m6A)” specificatamente nel 5′ UTR degli mRNA regola la normale fisiologia cellulare, la risposta infiammatoria e la progressione del cancro.Il ruolo e i meccanismi di m6A nelle malattie umane sono estesi ed è trattato in modo eccellente in altre revisioni complete (Yang et al., 2020; Knuckles e Bühler, 2018).
Primo tra questi, la vaccinazione molecolare SARS-CoV-2 induce condizioni di stress cellulare, come descritto dall’elevata segnalazione di NF-κB dopo la vaccinazione(Liu et al., 2021; Koo et al., 2010). In condizioni di stress cellulare, che può essere indotto da un’infezione virale o da stati patologici come il cancro, m6A media gli mRNA affinché subiscano la traduzione preferenzialmente in modo indipendente dal capside (Meyer et al., 2015). Come discusso in precedenza, questo è l’opposto dell’impatto della vaccinazione mRNA SARS-CoV-2, che spinge le cellule verso una traduzione cap-dipendente.Inoltre, in condizioni diversificate di stress cellulare, vi è un’induzione schiacciante dell’aggiunta di m6A a tutto il trascrittoma che fa sì che un numero maggiore di mRNA possieda 5’UTR arricchiti con m6A (Meyer et al., 2015).
Il fattore di inizio della traduzione eucariotica 4E (eIF4E) è la proteina legante il cappuccio iniziale dell’mRNA che dirige i ribosomi alla struttura del cappuccio degli mRNA, al fine di avviare la traduzione in proteine. La dipendenza dalla traduzione cap-dipendente degli mRNA del vaccino consumerà un surplus di disponibilità di eIF4E necessaria per tradurre un numero innaturalmente elevato di mRNA sintetici. Tuttavia, la traduzione indipendente dal limite avviene senza richiedere che eIF4E sia associato a eIF4F. La competizione per i ribosomi si sposterà verso la traduzione dei trascritti cap-indipendente, poiché gli mRNA sottoposti a traduzione cap-indipendente sono dotati, a parte i siti di ingresso dei ribosomi interni (IRES), di speciali motivi di legame che si legano a fattori che reclutano attivamente mRNA ai potenziatori traslazionali indipendenti dal cappuccio del ribosoma (CITE) (Shatsky et al., 2018). Inoltre, ciò significa anche che eIF4E, che è un potente regolatore oncogenico e modulatore della proliferazione cellulare, sosterrà le sue attività grazie a questa competizione per un periodo di tempo innaturalmente prolungato, cercando di controbilanciare la competizione tra mRNA con capside resistente nei vaccini e contenenti IRES mRNA(Kyriakopoulos e McCullough, 2021; Svitkin et al., 2005). Questo tipo di condizione si traduce in una disregolazione delle modifiche co-trascrizionali dell’mRNA m6A e si collega seriamente alle progressioni molecolari di vari tumori (Han e Choe, 2020), oltre a creare condizioni predisponenti per successive infezioni virali (Svitkin et al., 2005). Successivamente consideriamo l’impatto della glicoproteina spike SARS-CoV-2 derivata dalla vaccinazione mRNA sul sistema IFN cellulare attraverso la massiccia produzione di esosomi.
Esosomi e microRNA
Un’importante rete di comunicazione tra le cellule è costituita dalle vescicole extracellulari(EV) che vengono costantemente rilasciate da una cellula e successivamente assorbite da un’altra cellula, che potrebbe trovarsi in un organo distante. Piccole vescicole note come esosomi, formate all’interno degli endosomi, sono di dimensioni simili ai virus e vengono rilasciate attraverso l’esocitosi nello spazio extracellulare per circolare successivamente in tutto il corpo (Yoshikawa et al., 2019). Gli esosomi possono fornire una collezione diversificata di molecole biologicamente attive, tra cui mRNA, microRNA (miRNA), proteine, e lipidi (Ratajczak e Ratajczak, 2016). Durante un’infezione virale, le cellule infette secernono grandi quantità di esosomi che fungono da rete di comunicazione tra le cellule per orchestrare la risposta all’infezione(Chahar et al., 2015). In uno sforzo collaborativo di un team di ricercatori dell’Arizona e del Connecticut, è stato scoperto che le persone vaccinate con i vaccini mRNA hanno acquisito esosomi circolanti contenenti la glicoproteina spike SARS-CoV-2 entro il giorno 14 successivo alla vaccinazione(Bansal et al., 2021 ).Hanno anche scoperto che non c’erano anticorpi circolanti contro la glicoproteina spike quattordici giorni dopo il primo vaccino. Dopo il secondo vaccino, tuttavia, il numero di esosomi circolanti contenenti glicoproteina spike è aumentato fino a un fattore 12. Inoltre, gli anticorpi sono comparsi per la prima volta il giorno 14. Gli esosomi presentavano glicoproteina spike sulla loro superficie, che, secondo gli autori, indicavano una produzione di anticorpi facilitata. Quando i topi sono stati esposti a esosomi derivati da persone vaccinate, hanno sviluppato anticorpi contro la glicoproteina spike. È interessante notare che, dopo l’espressione di picco, il numero di esosomi circolanti contenenti glicoproteina spike è diminuito nel tempo, di pari passo con la diminuzione del livello di anticorpi contro la glicoproteina spike.
Biogenesis of extracellular vescicles from eukaryotic cell
Gli esosomi esistono come parte del meccanismo di decadimento dell’mRNA in stretta associazione in condizioni di stress con granuli di stress (SG) e corpi P (PB) (Decker e Parker, 2012; Kothandan et al., 2020). In condizioni di traduzione indotta da vaccino-mRNA, che potrebbe essere definita “eccessiva dipendenza dalla traduzione cap-dipendente”, esiste un’evidente resistenza alla promozione e all’assemblaggio del grande complesso di decapping (Kyriakopoulos e McCullough, 2021), e quindi resistenza contro processi fisiologici di decadimento dell’mRNA (Decker e Parker, 2012).Ciò significherebbe che il destino di particolari mRNA sintetici che altrimenti sarebbero determinati dalla strategia cellulare comune per il turnover dell’mRNA che coinvolge le ribonucleinproteine messaggere (mRNP) viene omesso (Borbolis e Syntichaki, 2015). Inoltre, in condizioni di eccessivo affidamento sulla traduzione cap-dipendente da parte degli mRNA sintetici nei vaccini SARS-CoV-2 (Kyriakopoulos e McCullough, 2021), molti mRNA nativi che contengono una considerevole IRES e metilazioni specifiche (m6A) nella loro struttura sceglieranno favorevolmente traduzione indipendente dal capside, che è fortemente legata ai meccanismi di controllo della qualità del decadimento dell’mRNA (Han e Choe, 2020). In questo senso, considerevoli prodotti di mRNA deadenilati così come prodotti derivati dal metabolismo dell’mRNA (decadimento) sono direttamente collegati ai carichi esosomici (Borbolis e Syntichaki, 2015). Un esempio di dipendenza dalla traduzione cap-dipendente è descritto nella leucemia linfoblastica acuta a cellule T (T-ALL). A causa del target meccanicistico della rapamicina C (mTORC) -1 sovrafunzionante in T-ALL, le cellule sono guidate completamente verso la traduzione cap-dipendente (Girardi e De Keersmaecker, 2015). Una condizione analoga è descritta da Kyriakopoulos e McCullough (2021). Anche in questo stato canceroso altamente aggressivo, durante l’inibizione della traduzione cap-dipendente nelle cellule T-ALL, c’è una rapida reversione alla traduzione cap-indipendente (Girardi e De Keersmaecker, 2015). Allo stesso modo, un’infezione da picornavirus (Jang et al., 1990) spinge le cellule verso la traduzione indipendente dal capside a causa dell’inibizione dei componenti del complesso eIF4F e del pluralismo di IRES nell’RNA virale.
Negli esseri umani, vi è un’abbondanza di infezioni da picornavirus per lo più asintomatiche come il virus Safford con una sieroprevalenza superiore al 90% nei bambini e negli adulti (Zoll et al., 2009). In entrambi i casi, sia che si tratti di un evento apoptotico dovuto a una condizione simile allo stress (Rusk, 2008) o di un effetto carcinomatoso simile a un mRNA-cap-driven (De Paolis et al., 2021), i livelli di miRNA saranno aumentati a causa del aumento del funzionamento epitrascrittomico e aumento del decadimento dell’mRNA. A causa dell’elevata domanda di espressione genica, saranno elevati livelli di alcuni miRNA dovrebbe essere contenuto negli esosomi tramite corpi P(Yu et al., 2016). Inoltre, in condizioni di produzione schiacciante di glicoproteina spike SARS-CoV-2 dovuta alla vaccinazione molecolare SARS-CoV-2, ci si aspetterebbe ovviamente che una percentuale significativa di glicoproteine spike intracellulari sovrabbondanti venga esportata anche tramite carichi esosomici ( Wei et al., 2021).Mishra e Banerjea (2021) hanno studiato il ruolo degli esosomi nella risposta cellulare delle cellule trasfettate con spike SARS-CoV-2. Hanno scritto nell’abstract:
“We propose that SARS-CoV-2 gene product, Spike, is able to modify the host exosomal cargo, which gets transported to distant uninfected tissues and organs and can initiate a catastrophic immune cascade within Central Nervous System (CNS).”
“Proponiamo che il prodotto del gene SARS-CoV-2, Spike, sia in grado di modificare il carico esosomico dell’ospite, che viene trasportato a tessuti e organi distanti non infetti e può avviare una cascata immunitaria catastrofica all’interno del sistema nervoso centrale (SNC).”
I loro esperimenti hanno coinvolto la crescita di cellule umane HEK293T in coltura e la loro esposizione ai plasmidi del gene spike SARS-CoV-2, che hanno indotto la sintesi della glicoproteina spike all’interno delle cellule. Hanno scoperto sperimentalmente che queste cellule rilasciavano abbondanti esosomi che ospitavano glicoproteina spike insieme a microRNA specifici.Hanno quindi raccolto gli esosomi e li hanno trasferiti in una coltura cellulare di microglia umana(le cellule immunitarie che risiedono nel cervello). Hanno dimostrato che la microglia assorbiva prontamente gli esosomi e rispondeva ai microRNA avviando una risposta infiammatoria acuta.Il ruolo della microglia nel causare la neuroinfiammazione in varie malattie virali, come il virus dell’immunodeficienza umana (HIV), il virus dell’encefalite giapponese(JEV)e la dengue, è ben consolidato.
Hanno proposto che la comunicazione cellula-cellula a lunga distanza tramite gli esosomi potrebbe essere il meccanismo mediante il quale i sintomi neurologici si manifestano nei casi gravi di COVID-19.
In un’ulteriore esplorazione, gli autori hanno identificato due microRNA che erano presenti in alte concentrazioni negli esosomi: miR-148a e miR-590. Hanno proposto un meccanismo specifico mediante il quale questi due microRNA interromperebbero in modo specifico la segnalazione dell’interferone di tipo I, attraverso la soppressione di due proteine critiche che controllano il percorso: la peptidasi 33 specifica dell’ubiquitina (USP33) e l’IRF9. Gli eterodimeri STAT1 e STAT2 fosforilati richiedono IRF9 per legare gli elementi di risposta stimolati da IFN, e quindi IRF9 svolge un ruolo essenziale nella risposta di segnalazione. Gli autori hanno dimostrato sperimentalmente che la microglia esposta agli esosomi estratti dalla coltura HEK293 aveva una diminuzione del 50% nell’espressione cellulare di USP33 e una diminuzione del 60% in IRF9. Hanno inoltre scoperto che miR-148a blocca specificamente USP33 e miR-590 blocca specificamente IRF9. USP33 rimuove l’ubiquitina da IRF9 e così facendo la protegge dal degrado. Pertanto, i due microRNA cospirano insieme per interferire con IRF9, bloccando così la risposta del recettore agli interferoni di tipo I. Uno studio di de Gonzalo-Calvo et al. (2021) hanno esaminato il profilo del microRNA nel sangue dei pazienti con COVID-19 e la loro varianza quantitativa basata sulla gravità della malattia. Si è scoperto che più miRNA erano regolati verso l’alto e verso il basso. Tra questi c’era miR-148a-3p, il filamento guida precursore di miR-148a. Tuttavia, il miR-148a stesso non era tra i microRNA catalogati come eccessivi o carenti nel loro studio, né lo era il miR-590. Da questi risultati risulta che miR148a e miR-590 e i loro effetti infiammatori sono unici per la produzione di glicoproteina spike SARS-CoV-2 indotta dalla vaccinazione. Studi con traccianti hanno dimostrato che, dopo l’iniezione nel muscolo del braccio, l’mRNA nei vaccini a mRNA viene trasportato nel sistema linfatico dalle cellule immunitarie e infine si accumula nella milza in alte concentrazioni (Bahl et al., 2017). Altri studi hanno dimostrato che le cellule immunitarie stressate nei centri germinali della milza rilasciano grandi quantità di esosomi che viaggiano verso i nuclei del tronco cerebrale lungo il nervo vago(come rivisto in Seneff e Nigh (2021)). Il nervo vago è il decimo nervo cranico ed entra nel tronco encefalico vicino alla laringe.
Il superiore e ricorrente i nervi laringei sono rami del vago che innervano le strutture coinvolte nella deglutizione e nel parlare. Le lesioni in questi nervi causano la paralisi delle corde vocali associata a difficoltà a deglutire (disfagia), difficoltà a parlare (disfonia) e/o mancanza di respiro (dispnea)(Gould et al., 2019; Erman et al., 2009). Torneremo su queste patologie specifiche nella nostra revisione dei dati VAERS di seguito. Le cellule HEK293 erano originariamente derivate da colture prelevate dal rene di un feto umano diversi decenni fa e immortalate attraverso l’infezione con il DNA dell’adenovirus. Sebbene siano state estratte dal rene, le cellule mostrano attraverso il loro profilo di espressione proteica che è probabile che siano di origine neuronale (Shaw et al., 2002).Ciò suggerisce che i neuroni nel nervo vago risponderebbero in modo simile alla glicoproteina spike SARS-CoV-2. Pertanto, le prove disponibili suggeriscono fortemente che la glicoproteina spike SARS-CoV-2 prodotta in modalità endogena crei un profilo di microRNA diverso rispetto all’infezione naturale con SARS-CoV-2 e tali differenze comportano una gamma potenzialmente ampia di effetti deleteri. Un punto centrale della nostra analisi di seguito è l’importante distinzione tra l’impatto della vaccinazione rispetto all’infezione naturale sull’IFN di tipo I. Mentre la vaccinazione sopprime attivamente la sua produzione, l’infezione naturale promuove la produzione di IFN di tipo I molto presto nel ciclo della malattia. Quelli con condizioni preesistenti spesso mostrano una segnalazione IFN di tipo I compromessa, che porta a COVID-19 più grave, critico e persino fatale. Se la compromissione indotta dal vaccino viene mantenuta mentre i livelli di anticorpi diminuiscono nel tempo, ciò potrebbe portare a una situazione in cui il vaccino provoca un’espressione della malattia più grave di quanto sarebbe avvenuto in assenza del vaccino. Un’altra conseguenza prevista della soppressione dell’IFN di tipo I sarebbe la riattivazione di infezioni virali croniche preesistenti, come descritto nellaSezione 9.
Riparazione del DNA compromessa e immunità adattativa
Il sistema immunitario e il sistema di riparazione del DNA sono i due sistemi primari su cui fanno affidamento gli organismi superiori per difendersi da diverse minacce e condividono elementi comuni.La perdita di funzione delle principali proteine di riparazione del DNA porta a difetti di riparazione che inibiscono la produzione di cellule B e T funzionali, con conseguente immunodeficienza.La riparazione dell’unione dell’estremità non omologa(NHEJ)svolge un ruolo fondamentale nella ricombinazione V (D) J specifica dei linfociti, che è essenziale per produrre il repertorio altamente diversificato di anticorpi delle cellule B in risposta all’esposizione all’antigene(Jiang e Mei, 2021) .
“La riparazione del DNA compromessa è anche una via diretta verso il cancro.“
Un articolo pubblicato daLiu et al., nel 2021 ha monitorato diversi parametri associati alla funzione immunitaria in una coorte di pazienti conducendo il sequenziamento dell’mRNA a cellula singola delle cellule mononucleari del sangue periferico (PBMC) raccolte dai pazienti prima e 28 giorni dopo la prima iniezione di un vaccino COVID-19 basato su una versione indebolita del virus(Liu et al., 2021). Sebbene questi vaccini siano diversi dai vaccini a mRNA, funzionano anche iniettando il contenuto del vaccino nel muscolo deltoide, aggirando le barriere mucose e vascolari. Gli autori hanno riscontrato un’alterazione consistente dell’espressione genica dopo la vaccinazione in molti diversi tipi di cellule immunitarie. Gli aumenti osservati nella segnalazione di NF-κB e le risposte IFN di tipo I ridotte sono stati ulteriormente confermati da test biologici. Coerentemente con altri studi, hanno scoperto che STAT2 e IRF7 erano significativamente sottoregolati 28 giorni dopo la vaccinazione, indicativi di risposte IFN di tipo I alterate. Hanno scritto: “Insieme, questi dati hanno suggerito che dopo la vaccinazione, almeno entro il giorno 28, oltre alla generazione di anticorpi neutralizzanti, il sistema immunitario delle persone, compresi quelli dei linfociti e dei monociti, era forse in uno stato più vulnerabile”. (Liu et al., 2021).
Questi autori hanno anche identificato cambiamenti inquietanti nell’espressione genica che implicherebbero una ridotta capacità di riparare il DNA. Fino al 60% dell’attività trascrizionale totale nelle cellule in crescita comporta la trascrizione del DNA ribosomiale (rDNA) per produrre RNA ribosomiale (rRNA). L’enzima che trascrive il DNA ribosomiale in RNA è l’RNA polimerasi I (Pol I). Pol I monitora anche l’integrità dell’rDNA e influenza la sopravvivenza cellulare (Kakarougkas et al., 2013). Durante la trascrizione, le RNA polimerasi (RNAP) scansionano attivamente il DNA per trovare lesioni voluminose (rotture a doppio filamento)e innescare la loro riparazione. Nelle cellule eucariotiche in crescita, la maggior parte della trascrizione comporta la sintesi dell’RNA ribosomiale da parte di Pol I. Pertanto, Pol I promuove la sopravvivenza in seguito a danni al DNA (Kakarougkas et al., 2013). Molti dei geni sottoregolati identificati da Liu et al. (2021) sono stati collegati al ciclo cellulare, al mantenimento dei telomeri e sia all’apertura del promotore che alla trascrizione di POL I, indicativi di processi di riparazione del DNA compromessi. Uno dei set di geni che sono stati soppressi era dovuto alla “deposizione di nuovo CENPA [proteina centromerica A] contenente nucleosomi al centromero”. Il CENPA di nuova sintesi è depositato nei nucleosomi al centromero durante la tarda telofase/la prima fase G1 del ciclo cellulare. Ciò indica l’arresto del ciclo cellulare nella fase G1 come caratteristica della risposta al vaccino SARS-CoV-2 inattivato. L’arresto di cellule staminali embrionali pluripotenti nella fase G1(prima dell’inizio della replicazione)comporterebbe un auto-rinnovamento compromesso e il mantenimento della pluripotenza(Choi et al., 2013). Due proteine checkpoint coinvolte in modo cruciale nella riparazione del DNA e nell’immunità adattativa sono BRCA1 e 53BP1, che facilitano sia la ricombinazione omologa (HR) che NHEJ, i due processi di riparazione primari (Zhang e Powell, 2005; Panier e Boulton, 2014). In un esperimento in vitro su cellule umane, è stato specificamente dimostrato che la glicoproteina spike a lunghezza intera SARS-CoV-2 entra nel nucleo e ostacola il reclutamento di queste due proteine di riparazione nel sito di una rottura del doppio filamento (Jiang e Mei, 2021). Gli autori hanno riassunto le loro scoperte dicendo: “Meccanicisticamente, abbiamo scoperto che la proteina spike si localizza nel nucleo e inibisce la riparazione del danno al DNA impedendo il reclutamento della proteina chiave di riparazione del DNA BRCA1 e 53BP1 nel sito del danno”.Un altro meccanismo attraverso il quale i vaccini a mRNA potrebbero interferire con la riparazione del DNA è attraverso il miR-148. Questo microRNA ha dimostrato di sottoregolare le risorse umane nella fase G1 del ciclo cellulare(Choi et al., 2014). Come accennato in precedenza in questo documento, questo era uno dei due microRNA trovati negli esosomi rilasciati dalle cellule umane in seguito alla sintesi della glicoproteina spike SARS-CoV-2 negli esperimenti di Mishra e Banerjea (2021).
A proposito di Trombocitopenia autoimmune
I vaccini a mRNA non sono privi di rischi per la VITT (trombocitopenia trombotica immunitaria indotta da vaccino), poiché sono stati pubblicati studi di casi che documentano tali eventi, inclusa la trombosi del seno venoso cerebrale mortale e fatale (vedi conclusioni della prima parte del lavoro in oggetto)(Lee et al., 2021; Akiyama et al., 2021; Atoui et al., 2022; Zakaria et al., 2021).Il meccanismo si ritiene che coinvolga gli anticorpi VITT che si legano al fattore piastrinico 4(PF4)e formino immunocomplessi che inducono l’attivazione piastrinica. Le successive cascate di coagulazione causano la formazione di microcoaguli diffusi nel cervello, nei polmoni, nel fegato, nelle gambe e altrove, associati a un drastico calo della conta piastrinica(Kelton et al., 2021). La reazione al vaccino è stata descritta come molto simile alla trombocitopenia indotta da eparina(HIT), tranne per il fatto che la somministrazione di eparina non è coinvolta(Cines and Bussel, 2021).È stato dimostrato che i vaccini mRNA suscitano principalmente una risposta immunitaria immunoglobulinica G(IgG), con quantità minori di IgA indotte(Wisnewski et al., 2021)e ancor meno produzione di IgM(Danese et al., 2021). La quantità di anticorpi IgG prodotti è paragonabile alla risposta osservata nei casi gravi di COVID-19. Sono gli anticorpi IgG in complesso con l’eparina che inducono l’HIT. Si può ipotizzare che le IgG complessate con la glicoproteina spike SARS-CoV-2 e PF4 siano il complesso che induce la VITT in risposta ai vaccini a mRNA. È stato infatti dimostrato sperimentalmente che il dominio di legame del recettore (RBD) della proteina spike si lega al PF4 (Passariello et al., 2021).
Il meccanismo alla base di HIT è stato ben studiato, anche attraverso l’uso di modelli murini umanizzati. È interessante notare che le piastrine umane, ma non le piastrine di topo, esprimono il recettore FcγRIIA, che risponde ai complessi PF4/eparina/IgG attraverso una cascata di fosforilazione della tirosina per indurre l’attivazione piastrinica. All’attivazione, le piastrine rilasciano granuli e generano microparticelle procoagulanti. Inoltre assorbono il calcio, attivano la proteina chinasi C, si aggregano in microtrombi e lanciano una cascata di morte cellulare tramite l’attivazione della calpaina. Queste piastrine attivate rilasciano PF4 nello spazio extracellulare, favorendo un circolo vizioso, poiché questo ulteriore PF4 si lega anche all’eparina e all’anticorpo IgG per promuovere ulteriormente l’attivazione piastrinica. Pertanto, FcγRIIA è fondamentale per il processo patologico(Nevzorova et al., 2019).
Studi su topi ingegnerizzati per esprimere il recettore FcγRIIA umano hanno dimostrato che questi topi transgenici sono molto più suscettibili alla trombocitopenia rispetto alle loro controparti wild type (McKenzie et al., 1999). È stato proposto che le piastrine possano svolgere un ruolo importante nella clearance dei complessi antigene-anticorpo intrappolando l’antigene nei trombi e/o trasportandolo nella milza per la rimozione da parte delle cellule immunitarie. Le piastrine vengono ovviamente rapidamente consumate nel processo, il che si traduce quindi in un basso numero di piastrine(trombocitopenia).
Le piastrine normalmente circolano con una vita media di soli cinque-nove giorni, quindi vengono costantemente sintetizzate nel midollo osseo e eliminate nella milza. Le piastrine legate agli anticorpi, dopo l’attivazione piastrinica tramite i recettori Fcγ, migrano verso la milza dove vengono intrappolate e rimosse attraverso la fagocitosi dai macrofagi (Crow e Lazarus, 2003). Completamente un terzo delle piastrine totali del corpo si trovano nella milza. Poiché i vaccini a mRNA vengono trasportati nella milza da cellule immunitarie inizialmente attratte dal sito di iniezione nel muscolo del braccio, esiste un’enorme opportunità per il rilascio di esosomi contenenti spike-glicoproteina da parte delle cellule dendritiche nella milza che sintetizzano la proteina spike. Si può ipotizzare che l’attivazione piastrinica in seguito alla formazione di un complesso proteico P4F/IgG/spike nella milza sia parte del meccanismo che tenta di eliminare la glicoproteina spike tossica.
Abbiamo accennato in precedenza che uno dei due microRNA altamente espressi negli esosomi rilasciati dalle cellule umane esposte alla glicoproteina spike SARS-CoV-2 era miR-148a. È stato dimostrato sperimentalmente che miR-148a sopprime l’espressione di una proteina che svolge un ruolo centrale nella regolazione dell’espressione di FcγRIIA sulle piastrine. Questa proteina, chiamata T-cell ubiquitin ligand-2 (TULA-2), inibisce specificamente l’attività del recettore piastrinico Fcγ. miR-148a prende di mira l’mRNA di TULA-2 e ne riduce l’espressione. Pertanto, miR-148a, presente negli esosomi rilasciati dai macrofagi che sono costretti dal vaccino a sintetizzare la glicoproteina spike SARS-CoV-2, agisce per aumentare il rischio di trombocitopenia in risposta agli immunocomplessi formati dall’antigene della glicoproteina spike e dagli anticorpi IgG prodotti contro la glicoproteina spike.
PPAR-α, sulfatide e malattie del fegato
Come abbiamo già affermato, un esperimento di Mishra e Banerjea(2021)ha dimostrato che la glicoproteina spike SARS-CoV-2 induce il rilascio di esosomi contenenti microRNA che interferiscono specificamente con la sintesi di IRF9. In questa sezione mostreremo che una delle conseguenze della soppressione di IRF9 sarebbe la ridotta sintesi di solfatide nel fegato, mediata dal recettore nucleare attivato dal proliferatore del perossisoma recettore α (PPAR-α).
I solfatidi sono i principali sfingoglicolipidi sierici dei mammiferi che sono sintetizzati e secreti principalmente dal fegato (Lu et al., 2019). Sono gli unici sfingolipidi solfonati nel corpo. I solfatidi sono formati da un processo in due fasi che comporta la conversione della ceramide in galattocerebroside e la sua successiva solfatazione. Il solfatide è espresso sulla superficie delle piastrine, degli eritrociti e dei linfociti. I solfatidi sierici esercitano funzioni sia anticoagulanti che anti-attivazione piastrinica. L’enzima nel fegato che sintetizza il solfatide, cerebroside sulfotransferasi, è stato specificamente trovato essere indotto dall’attivazione di PPAR-α nei topi (Kimura et al., 2012). Pertanto, la ridotta espressione di PPAR-α porta alla carenza di solfatide.
I ligandi PPAR-α mostrano effetti antinfiammatori e antifibrotici, mentre la carenza di PPAR-α porta a steatosi epatica, steatoepatite, steatofibrosi e cancro al fegato (Wang et al., 2020b). Nel 2019, un team di ricercatori in Giappone ha condotto un esperimento su topi con un gene difettoso per PPAR-α (Lu et al., 2019). Questi topi, se nutriti con una dieta ricca di colesterolo, erano suscettibili all’accumulo eccessivo di trigliceridi e all’esacerbazione dell’infiammazione e dello stress ossidativo nel fegato, insieme a livelli aumentati di fattori della coagulazione. I topi hanno anche manifestato livelli ridotti di solfatidi sia nel fegato che nel siero. Gli autori hanno ipotizzato che il sovraccarico di colesterolo eserciti i suoi effetti tossici in parte aumentando la trombosi, a seguito di un metabolismo lipidico epatico anormale e di stress ossidativo. Hanno dimostrato che il PPAR-α può attenuare questi effetti tossici attraverso la regolazione trascrizionale dei fattori della coagulazione e la sovraregolazione della sintesi del solfatide, oltre ai suoi effetti nel migliorare le malattie del fegato. Hanno proposto che terapie come i fibrati volti ad attivare il PPAR-α potrebbero prevenire le malattie cardiovascolari indotte da una dieta ad alto contenuto di colesterolo.
Studi con traccianti hanno dimostrato che l’mRNA dei vaccini a mRNA migra preferenzialmente verso il fegato e la milza, raggiungendo concentrazioni più elevate rispetto a qualsiasi altro organo ( Bahl et al., 2017 ). Pertanto, esiste il potenziale per la soppressione dell’IRF9 nel fegato da parte del vaccino. IRF9 è altamente espresso negli epatociti, dove interagisce con PPAR-α, attivando i geni bersaglio PPAR-α. Uno studio sui topi knockout IRF9 ha mostrato che questi topi hanno sviluppato steatosi e insulino-resistenza epatica se esposti a una dieta ricca di grassi. Al contrario, la sovraespressione epatica di IRF9 mediata da adenovirus nei topi obesi ha migliorato la sensibilità all’insulina e migliorato la steatosi e l’infiammazione ( Wang et al., 2013 ).
Numerosi casi clinici nella letteratura di ricerca descrivono danni al fegato in seguito a vaccini a mRNA ( Zin Tun et al., 2021 ; Dumortiera, 2022 ; Mann et al., 2021 ). Un fattore plausibile che porta a questi risultati è la soppressione del PPAR-α attraverso la downregulation dell’IRF9 e successivamente la diminuzione della sintesi del solfatide nel fegato.
Sindrome di Guillain Barré e sindromi da danno neurologico
La GBS è una neuropatia infiammatoria demielinizzante acuta associata a morbilità di lunga durata e un significativo rischio di mortalità ( Cr é ange, 2000 ). La malattia comporta un attacco autoimmune ai nervi associato al rilascio di citochine pro-infiammatorie.
Il GBS è spesso associato ad autoanticorpi contro il solfatide e altri sfingolipidi ( Ilyas et al., 1991 ). Le cellule T attivate producono citochine in risposta alla presentazione dell’antigene da parte dei macrofagi e queste citochine possono indurre la produzione di autoanticorpi attraverso la diffusione dell’epitopo ( Vanderlugt e Miller, 2002 ). Gli anticorpi, a loro volta, inducono l’attivazione del complemento, che causa demielinizzazione e danno assonale, portando a gravi lesioni ai neuroni periferici ( Kuwahara e Kusunoki, 2018 ). È stato dimostrato che la glicoproteina spike SARS-CoV-2 si lega all’eparan solfato, che è un complesso ammino-zuccherino solfato simile al galattosio solfatato nel solfatide ( Kalra e Kandimalla, 2021). Pertanto, è concepibile che la glicoproteina spike si leghi anche al sulfatide e ciò potrebbe innescare una reazione immunitaria al complesso spike-glicoproteina-sulfatide.
Come descritto nella sezione precedente, la ridotta sintesi di solfatidi nel fegato dovuta alla soppressione dell’IRF9 porterà nel tempo a una carenza sistemica di solfatidi. La carenza di solfatidi può avere un impatto importante sul cervello e sul sistema nervoso. Il 20% dei galattolipidi presenti nella guaina mielinica sono solfatidi. Il solfatide è un componente importante del sistema nervoso, presente in concentrazioni particolarmente elevate nella guaina mielinica sia nel sistema nervoso periferico che in quello centrale. Le carenze di sulfatide possono portare a debolezza muscolare, tremori e atassia ( Honke, 2013 ), che sono sintomi comuni di GBS. La neuroinfiammazione cronica mediata dalla microglia e dagli astrociti nel cervello porta a drammatiche perdite di sulfatide cerebrale e le carenze cerebrali di sulfatide sono una delle principali caratteristiche della malattia di Alzheimer.Qiu et al., 2021 ). I topi con un difetto nella capacità di sintetizzare il solfatide dalla ceramide mostrano una ridotta capacità di mantenere la salute degli assoni mentre invecchiano. Nel corso del tempo, sviluppano guaine mieliniche ridondanti, non compattate e in degenerazione, nonché una struttura deteriorante nei nodi di Ranvier negli assoni, causando la perdita di una giunzione assogliale funzionalmente competente ( Marcus et al., 2006 ).
L’angiotensina II (Ang II), oltre ai suoi profondi effetti sulle malattie cardiovascolari, svolge anche un ruolo nell’infiammazione nel cervello che porta alla malattia neurodegenerativa ( Lanz. et al., 2010 ). La glicoproteina spike SARS-CoV-2 contiene un sito di clivaggio della furina unico non trovato in SARS-CoV, che consente all’enzima extracellulare furin di staccare il segmento S1 della glicoproteina spike e rilasciarlo in circolazione ( Letarov et al., 2021 ) . È stato dimostrato che S1 attraversa la barriera emato-encefalica nei topi ( Rhea et al., 2021). S1 contiene il dominio di legame del recettore che si lega ai recettori ACE2, disabilitandoli. Quando la segnalazione del ricevitore ACE2 è ridotta, la sintesi di Ang II è aumentata. I neuroni nel cervello possiedono recettori ACE2 che sarebbero suscettibili di interruzione da parte di S1 rilasciato da esosomi contenenti spike-glicoproteina o cellule produttrici di spike-glicoproteina che avevano assorbito le nanoparticelle nei vaccini. Ang II migliora la segnalazione mediata da TLR4 nella microglia, inducendo l’attivazione della microglia e aumentando la produzione di specie reattive dell’ossigeno che portano al danno tissutale, all’interno del nucleo paraventricolare nel cervello ( Rodriguez-Perez et al., 2015 ).
Livelli elevati di Ang II sono un fattore causale nella neurodegenerazione del nervo ottico, causando neurite ottica, che può provocare una grave perdita irreversibile della vista ( Guo et al., 2017 ). Numerosi casi clinici hanno descritto casi di neuropatia ottica che compaiono poco dopo la vaccinazione mRNA per COVID-19 ( Maleki, 2021 ; Barone et al., 2021). Poco dopo la vaccinazione compaiono anche altre condizioni neurologiche debilitanti, in cui si sospetta una relazione causale. Un caso di studio con sede in Europa che ha monitorato i sintomi neurologici dopo la vaccinazione COVID-19 ha identificato 21 casi che si sono sviluppati entro una mediana di 11 giorni dopo la vaccinazione. I casi avevano diagnosi diverse tra cui trombosi del seno venoso cerebrale, malattie demielinizzanti del sistema nervoso, neuropatie periferiche infiammatorie, miosite, miastenia, encefalite limbica e arterite a cellule giganti ( Kaulen et al., 2021 ). Khayat-Khoei et al. (2021)descrivono una serie di casi di 7 pazienti, di età compresa tra 24 e 64 anni, che presentano malattia demielinizzante entro 21 giorni dalla prima o dalla seconda vaccinazione con mRNA. Quattro avevano una precedente storia di SM (controllata), mentre tre erano precedentemente sani.
La perdita dell’udito e l’acufene sono anche effetti collaterali ben noti di COVID-19. Un caso di studio ha coinvolto una serie di dieci pazienti COVID-19 che soffrivano di sintomi audiovestibolari come perdita dell’udito, disfunzione vestibolare e tinnito ( Jeong et al., 2021 ). Gli autori hanno dimostrato che il tessuto dell’orecchio interno umano esprime ACE2, furina e la proteasi transmembrana serina 2 (TMPRSS2), che facilita l’ingresso virale. Hanno anche dimostrato che SARS-CoV-2 può infettare specifici tipi di cellule dell’orecchio interno umano.
Un altro studio che ha valutato il potenziale del virus SARS-CoV-2 di infettare l’orecchio ha esaminato specificamente l’espressione del recettore ACE2 e degli enzimi furina e TM-PRSS2 di vari tipi di cellule nell’orecchio medio e interno dei topi. Hanno scoperto che ACE2 e furina erano “diffusamente presenti nella tromba di Eustachio, negli spazi dell’orecchio medio e nella coclea, suggerendo che questi tessuti sono suscettibili all’infezione da SARS-CoV-2”. ( Uranaka et al., 2021 ). L’acufene è positivamente associato all’ipertensione, che è indotta da livelli elevati di Ang II ( Rodrigues Figueiredo et al., 2016 ).
Il mal di testa è una reazione avversa molto comune ai vaccini mRNA COVID-19, in particolare per le persone che sono già suscettibili al mal di testa. In uno studio basato su un questionario che ha coinvolto 171 partecipanti, l’incidenza del mal di testa è risultata essere del 20,5% dopo il primo vaccino, salendo al 45,6% dopo il secondo colpo (Sekiguchi et al., 2021 ) . Un caso di studio ha descritto una donna di 37 anni che soffriva di un debilitante attacco di emicrania della durata di 11 giorni dopo il secondo vaccino mRNA Pfizer/BioNtech (Consoli et al ., 2021 ).
Gli steroidi sono spesso usati come terapia aggiuntiva per trattare l’emicrania ( Huang et al., 2013 ). Il desametasone e altri steroidi stimolano i recettori PPAR-α nel fegato attraverso il recettore degli steroidi, compensando così gli effetti della soppressione dell’IRF9 ( Lemberger et al., 1994 ). Una teoria per le origini dell’emicrania coinvolge l’elaborazione alterata dell’input sensoriale nel tronco cerebrale, principalmente nei neuroni del trigemino ( Dodick e Silberstein, 2006). Il nervo trigemino si trova in prossimità del nervo vago nel tronco encefalico, quindi gli esosomi che trasportano glicoproteine a punta potrebbero facilmente raggiungerlo lungo la via vagale. La risonanza magnetica ha rivelato che i cambiamenti strutturali nel nervo trigemino che riflettono la microstruttura aberrante e la demielinizzazione sono una caratteristica delle persone che soffrono di frequenti emicranie ( Mungoven et al., 2020 ). Un potenziale fattore legato all’infezione da SARS-CoV-2 o alla vaccinazione con mRNA è un livello eccessivo di Ang II nel tronco encefalico a causa dell’inibizione della glicoproteina spike SARS-CoV-2 dei recettori ACE2. Gli ACE-inibitori e gli antagonisti del recettore Ang II sono diventati farmaci popolari per il trattamento dell’emicrania off-label ( Tronvik et al., 2003 ; Nandha e Singh, 2012). L’emicrania potrebbe quindi derivare sia dall’interruzione dei recettori ACE2 da parte della glicoproteina spike sia dalla distruzione della guaina mielinica che copre i nervi facciali critici attraverso una risposta infiammatoria della microglia e la perdita di sulfatide. La fonte di quella glicoproteina spike potrebbe essere esogena o endogena.
Paralisi di Bell
La paralisi di Bell è una comune neuropatia cranica che causa una paralisi facciale unilaterale. Anche negli studi clinici di fase III, la paralisi di Bell si è distinta, con sette casi comparsi nel braccio di trattamento rispetto a un solo caso nel gruppo placebo ( FDA, 2021a ; FDA, 2021b ). Un caso di studio riportato in letteratura ha coinvolto un uomo di 36 anni che ha sviluppato debolezza al braccio sinistro un giorno dopo la vaccinazione, progredendo in intorpidimento e formicolio al braccio e conseguenti sintomi della paralisi di Bell nei giorni successivi. Una causa comune della paralisi di Bell è la riattivazione dell’infezione da virus herpes simplex centrata attorno al ganglio genicolato ( Eviston et al., 2015 ). Questo, a sua volta, può essere causato dall’interruzione della segnalazione IFN di tipo I.
Miocardite
C’è stata una notevole attenzione da parte dei media dedicata al fatto che i vaccini COVID-19 causano miocardite e pericardite, con un aumento del rischio in particolare per gli uomini di età inferiore ai 50 anni ( Simone et al., 2021 ; Jain et al., 2021 ). È stato dimostrato che la glicoproteina spike SARS-CoV-2 danneggia i periciti cardiaci, che supportano i capillari e i cardiomiociti ( Avolio et al., 2020 ). La miocardite è associata all’attivazione piastrinica, quindi questo potrebbe essere un fattore in gioco nella risposta ai vaccini ( Weikert. et al., 2002 ). Tuttavia, un altro fattore potrebbe essere correlato agli esosomi rilasciati dai macrofagi che hanno assorbito le nanoparticelle di mRNA e i microRNA specifici trovati in quegli esosomi.
Uno studio che ha coinvolto pazienti affetti da grave malattia COVID-19 ha esaminato specificamente l’espressione dei microRNA circolanti rispetto ai pazienti affetti da influenza e ai controlli sani. Un microRNA che è stato costantemente sovraregolato in associazione con COVID-19 era il miR-155 e gli autori hanno suggerito che potrebbe essere un predittore di danno miocardico cronico e infiammazione. Al contrario, l’infezione influenzale non era associata ad una maggiore espressione di miR-155. Hanno concluso: “Il nostro studio ha identificato livelli significativamente alterati di miR [microRNA] associati al cuore nei pazienti con COVID-19, indicando una forte associazione di COVID-19 con disturbi cardiovascolari e rispettivi biomarcatori” ( Garg et al., 2021 ) .
Uno studio che ha confrontato 300 pazienti con malattie cardiovascolari con controlli sani ha mostrato un aumento statisticamente significativo dei livelli circolanti di miR-155 nei pazienti rispetto ai controlli. Inoltre, quelli con arterie più ristrette (secondo un punteggio Gensini) avevano livelli più alti rispetto a quelli con malattia minore ( Qiu e Ma, 2018 ).
È importante sottolineare che gli esosomi svolgono un ruolo nell’infiammazione in associazione con le malattie cardiache. Durante l’infarto del miocardio, il miR-155 è nettamente sovraregolato nei macrofagi nel muscolo cardiaco e rilasciato nell’ambiente extracellulare all’interno degli esosomi. Questi esosomi vengono consegnati ai fibroblasti e il miR-155 sottoregola le proteine nei fibroblasti che proteggono dall’infiammazione e promuovono la proliferazione dei fibroblasti. La compromissione risultante porta alla rottura cardiaca ( Wang et al., 2017b ).
Abbiamo già discusso di come il segmento S1 della glicoproteina spike SARS-CoV-2 possa essere scisso dalla furina e rilasciato in circolazione. Si lega ai recettori ACE2 attraverso il suo dominio di legame del recettore (RBD) e questo inibisce la loro funzione. Poiché l’ACE2 degrada l’Ang II, la disattivazione dell’ACE2 porta direttamente alla sovraespressione dell’Ang II, aumentando ulteriormente il rischio di malattie cardiovascolari. La vasocostrizione indotta da AngII è un meccanismo indipendente per indurre un danno miocardico permanente anche quando non è presente un’ostruzione coronarica. Episodi ripetuti di costrizione improvvisa di un’arteria cardiaca dovuti all’Ang II possono alla fine portare a insufficienza cardiaca o morte improvvisa ( Gavras e Gavras, 2002 ). Sono stati descritti casi fatali di vaccinazione COVID-19 ( Choi et al., 2021 ; Verma et al., 2021).
La soppressione di ACE2 era già stata osservata negli studi sul virus SARS-CoV originale. Uno studio autoptico su pazienti che soccombono alla SARS-CoV ha rivelato un ruolo importante per l’inibizione dell’ACE2 nel promuovere il danno cardiaco. L’RNA virale SARS-CoV è stato rilevato nel 35% di 20 campioni di cuore umano sottoposti ad autopsia prelevati da pazienti deceduti. C’è stato un marcato aumento dell’infiltrazione di macrofagi associata a danno miocardico nei pazienti i cui cuori erano stati infettati da SARS-CoV. È importante sottolineare che la presenza di SARS-CoV nel cuore è stata associata a una marcata riduzione dell’espressione della proteina ACE2 ( Oudit et al., 2009 ).
Alcuni esempi di come viene spiegata la “sicurezza” dei vaccini al grande pubblico dalle corporations del farmaco e del come delle certezze scientifiche sbandierate per molti decenni, scompaiono letteralmente dalla rete perché sinceramente “scomode”. Ma io le ho recuperate con la “Waybackmachine”…
In che modo la FDA valuta la sicurezza dei vaccini?
“I vaccini vengono sottoposti a test rigorosi e approfonditi per determinarne la sicurezza e l’efficacia. Scienziati altamente qualificati e personale medico della FDA esaminano attentamente tutte le informazioni in una domanda di marketing prima che un vaccino possa essere approvato per l’uso da parte del pubblico. Dopo l’approvazione, la FDA controlla attentamente anche la qualità dei vaccini: tutti i lotti prodotti devono superare i test prima di poter essere utilizzati. E come con tutti i produttori di farmaci, i produttori di vaccini devono seguire rigorosi standard di produzione. Inoltre, la FDA conduce ispezioni di routine dei siti di produzione. La FDA lavora anche a stretto contatto con i Centers for Disease Control and Prevention (CDC) per monitorare le segnalazioni di effetti collaterali (eventi avversi) dei vaccini. FDA e CDC prendono sul serio tutti i rapporti e collaborano per valutare e affrontare eventuali problemi potenziali”. (Food and Drug Administration – FDA May, 31 2016) – Documento cancellato dal sito ufficiale della FDA ma recuperato attraverso la waybackmachine a questo link: https://web.archive.org/web/20170528213949/https://www.fda.gov/AboutFDA/Transparency/Basics/ucm194586.htm
Sicurezza dei vaccini
“La ricerca e la sperimentazione sulla sicurezza sono alla base di ogni aspetto dello sviluppo e della produzione di vaccini in Australia. Prima che i vaccini siano resi disponibili per l’uso, vengono rigorosamente testati su migliaia di persone in studi clinici progressivamente più grandi che sono rigorosamente monitorati per la sicurezza. I risultati di questi studi costituiscono la base per un processo continuo di test e monitoraggio che dura per tutta la vita di ciascun vaccino. Tutti i vaccini registrati in Australia dalla Therapeutical Goods Administration (TGA) vengono valutati per garantire che siano efficaci, conformi a rigorosi standard di produzione e che abbiano un solido record di sicurezza. Ciò include test rigorosi per ciascun componente del vaccino, inclusi conservanti, additivi e adiuvanti del vaccino. Possono essere necessari fino a 10 anni prima che un vaccino venga approvato per l’uso”. (Australian Government, Department of Health, Immunise Australia Program. April 20, 2015) Documento cancellato dal sito ufficiale della FDA ma recuperato attraverso la waybackmachine a questo link : https://web.archive.org/web/20150426041949/http://www.immunise.health.gov.au/internet/immunise/publishing.nsf/Content/safety-of-vaccines
“I vaccini sono tenuti ai più alti standard di sicurezza. Gli Stati Uniti hanno attualmente la fornitura di vaccini più sicura ed efficace della storia. I vaccini sono sottoposti a un rigoroso ed ampio programma di valutazione per determinare la sicurezza e l’efficacia. Se il vaccino riceve l’approvazione dalla FDA, viene continuamente monitorato per la sicurezza e l’efficacia”. (U.S. Department of Health & Human Services accessed on January, 28 2017). Originariamente qui, (poi eliminato dal sito ufficiale del governo USA): https://www.vaccines.gov/basics/safety/informed/ adesso visibile qui: https://web.archive.org/web/20150426041949/http://www.immunise.health.gov.au/internet/immunise/publishing.nsf/Content/safety-of-vaccines
“Osservare i bambini vaccinati per molti anni alla ricerca di condizioni di salute a lungo termine non sarebbe pratico e non sarebbe etico negare ai bambini un vaccino efficace mentre sono in corso studi a lungo termine”. (CDC Centers for Disease Control and Prevention – CDC: Parent’s Guide to childhood immunization. August 2015 p.33) https://www.cdc.gov/vaccines/parents/tools/parents-guide/downloads/parents-guide-508.pdf
Le affermazioni secondo cui i vaccini “vengono sottoposti a test rigorosi e approfonditi”, che sono “mantenuti secondo i più alti standard di sicurezza”, ecc., sono bugie. Gli studi che sono alla base delle autorizzazioni all’immissione in commercio sono un completo e totale “pasticcio metodologico”.Sono progettati così male che non consentono il rilevamento e la valutazione degli effetti della vaccinazione a breve termine, figuriamoci a lungo termine. Tali studi metodologicamente così inadeguati non dovrebbero mai far parte del processo di rilascio delle autorizzazioni all’immissione in commercio. Devo sottolineare qui che l’utilizzo di massa di vaccini non testati, cioè vaccini che non sono stati testati in modo affidabile, non è raro. Al contrario, la concessione di un’autorizzazione all’immissione in commercio per i vaccini senza studi metodologicamente validi di sicurezza ed efficacia è la regola, non un’eccezione.
La patogenesi del diabete di tipo 1 è complessa ed è il risultato di una combinazione di fattori genetici, ambientali, ormonali e immunologici. I fattori ambientali includono virus patogeni, dieta, tossine e stress così come i vaccini (Tishler and Shoenfeld, 2004). La possibile associazione tra T1D e i vaccini è ancora in discussione. Numerosi studi hanno cercato collegamenti tra diabete e vaccinazioni. Classen ha trovato che se la prima vaccinazione nei bambini viene eseguita dopo i 2 mesi di età, c’è un aumento del rischio di diabete (Classen, 1996). “Vaccinations and Type 1 Diabetes” – Alessandro Antonelli, Silvia Martina Ferrari, Andrea Di Domenicantonio, Ele Ferrannini, and Poupak Fallahi – Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy Pubblicato al capitolo 25 del volume “Vaccines & Autoimmunity” by Yehuda Shoenfeld et al 2015 – Ed. Wiley.
Il Lupus eritematoso è un’altra patologia autoimmune sulla quale vi sono studi che evidenziano una sua correlazione con le vaccinazioni: (capitolo 22) “Systemic Lupus Erythematosus Induced by Vaccines” – Nurit Katz-Agranov and Gisele Zandman-Goddard del volume “Vaccines & Autoimmunity” by Yehuda Shoenfeld et al 2015 – Ed. Wiley. Numerosi reports e studi hanno provato a collegare le manifestazioni del lupus sistemico eritematoso con il vaccino antiepatite B (HBV) ma essi hanno mostrato solo una correlazione temporale (Older et al., 1999).
Con riferimento agli studi sugli effetti del vaccino quadrivalente Gardasil, un’attenta valutazione del rischio individualizzato dei pazienti portatori di una storia medica di malattie autoimmuni e infettive(talora in forma non evidente)e di reazioni avverse alle precedenti vaccinazioni è richiesta(Bijl et al., 2012), oltre alla valutazione successiva ad ogni richiamo vaccinale (Agmon-Levin et al., 2009a, b).
Fattori di rischio per le patologie autoimmuni
Quanto alle disfunzioni tiroidee(non poche delle quali hanno una natura autoimmune), la letteratura scientifica e l’esperienza clinica mostrano che i metalli svolgono un ruolo chiave nello sviluppo di malattie autoimmuni (Hybenova,M., Hrda, P., Procházková, J., et al. (2010). The role of environmental factors in autoimmune thyroiditis. Neuro Endocrinol Lett, 31(3): 283–9.).
Questi sono frammenti di una letteratura scientifica che pongono dei seri dubbi sull’innocuità dei vaccini, soprattutto alla luce del fatto che non esistono studi che ne convalidino la prevalenza dei benefici a fronte dei rischi.
Allo stato attuale, oltre ad essere sconosciuto il vero fisiologismo di azione dei vaccini, è ignota la risposta del sistema immunitario dinnanzi ad un’immunizzazione che non è permanente ma che necessita di richiami.
Con tutti i dubbi esistenti sull’efficacia ed efficienza e la sicurezza.
A fronte di tutto questo, la presenza di patologie familiari (tra l’altro su base autoimmune), soprattutto davanti all’assenza di cautele su base individuale e alla totale ignoranza circa l’esistenza di tali peculiarità da parte dei medici, deve sconsigliare fortemente l’adozione della pratica vaccinale.
Esiste ormai una notevole letteratura scientifica sui danni da vaccini (praticamente tutti quelli per uso pediatrico) e il personale sanitario è autorizzato legalmente a non prenderli assolutamente in considerazione. Sperare di ottenere un’esonero è pertanto una pia illusione. La scelta di non vaccinare appartiene alla sua consapevolezza genitoriale e nessuno può scardinarla.
La valutazione degli HLA permetterebbe di denotare a grandi linee la presenza di immunodepressione ma resterebbero sempre decine (se non diverse centinaia) di valutazioni (polimorfismi genetici) a cui si devono sommare quelli ancora da scoprire e le loro implicazioni con il funzionamento del sistema immunitario. Uno di questi è il test MTHFR (vedi articolo: MTHFR & Disturbi Neurologici Indotti dalle Vaccinazioni). Ma non è l’unico.
In linea di principio, tutto ciò che non è dimostratamente innocuo è potenzialmente dannoso. Il principio di precauzione è sacro.
Stimolazione di autoanticorpi post vaccinali indotta da vaccinazione in soggetti apparentemente sani.
Negli studi evidenziati negli screenshots, un riferimento scientifico sulla possibilità – accertata clinicamente – che tale induzione alla produzione autoanticorpale avvenga, oltre che in soggetti sani (primo screen), anche in soggetti portatori di patologie autoimmuni (secondo screen). Questi screen sono tratti da “Vaccines & Autoimmunity” del Dr Yehuda Shoenfeld et al. Wiley Editions, 2015. Nel primo screen si legge:
“….L’induzione di autoanticorpi è stata studiata in bambini sani successivamente a vaccinazione con vaccini antiepatite A e antiepatite B. L’induzione di autoanticorpi 6 anni dopo l’inoculazione del vaccino antiepatite B è stata studiata in 210 bambini immunizzati alla nascita con il vaccino antiepatite B ricombinante (Belloni et al., 2010)…”
Nel secondo screen una tabella in cui sono riportati i dati relativi agli autoanticorpi sviluppati in soggetti portatori di patologie autoimmuni a seguito di vaccinazioni antiinfluenzale, antipneumococcica e antipapilloma virus. Ragioni a pieno sostegno del rifiuto totale di qualsiasi vaccinazione.
Ulteriori conferme sui danni indotti dai vaccini mediante produzione di autoanticorpi:
“…Come negli esseri umani, l’induzione di autoanticorpi è possibile in seguito alla vaccinazione negli animali. Gli autoanticorpi sono più frequentemente presenti nei cani più vecchi che in quelli più giovani (Papini et al., 2005). Di conseguenza, quando i cani anziani vengono vaccinati e testati per gli autoanticorpi dopo la vaccinazione, è difficile definire quale sia una conseguenza diretta del vaccino e cosa si è accumulato durante la vita. Nel primo studio sperimentale controllato per studiare la produzione di autoanticorpi dopo la vaccinazione di routine, i ricercatori hanno arruolato giovani cani che non erano stati precedentemente vaccinati (Hogenesch et al., 1999).”
“…Hanno dimostrato che la vaccinazione obbligatoria contro la rabbia, il virus del cimurro canino e il parvovirus canino 9 ha provocato la produzione di svariati autoanticorpi…”
Autoanticorpi indotti dalle vaccinazioni nei cani
Già nel 2009 Toplak e Avcin avevano ipotizzato la produzione autoanticorpale indotta dai vaccini e la loro responsabilità nell’induzione di patologie autoimmuni.
Leggete che cosa concludevano:
“…La vaccinazione contro i microrganismi patogeni è uno dei principali risultati della medicina moderna, ma a causa di un numero crescente di segnalazioni di reazioni avverse la procedura di vaccinazione ha indotto anche un considerevole dibattito. È noto che alcune infezioni sono coinvolte nell’attivazione della produzione di autoanticorpi, che potrebbero portare a reazioni avverse autoimmuni in soggetti geneticamente predisposti.
Sulla base di questi risultati è stato ipotizzato che le vaccinazioni potrebbero indurre reazioni autoimmuni simili.
Allo stato attuale (ndr: era il 2009) non vi è alcuna prova evidente che le vaccinazioni siano associate a malattie autoimmuni conclamate, ma
è stato dimostrato che nelle persone geneticamente predisposte la vaccinazione potrebbe innescare la produzione di autoanticorpi e reazioni avverse autoimmuni.
I primi studi che osservavano la produzione di autoanticorpi dopo la vaccinazione sono stati condotti su cani e topi.
Diversi studi hanno successivamente dimostrato la produzione di autoanticorpi dopo la vaccinazione in pazienti con malattie autoimmuni,
ma ci sono solo dati limitati sulle risposte autoimmuni dopo le vaccinazioni in esseri umani apparentemente sani (ndr: il Dr Yehuda Shoenfeld confermerà nel 2015). Questa recensione riassume le prove attuali (ndr: anno 2009) sugli autoanticorpi indotti da vaccinazione in soggetti apparentemente sani, compresi gli studi su animali ed esseri umani.
Vuol dire reazioni e patologie autoimmuni presto e/o più avanti negli anni.. (Autoantibodies Induced by Vaccine – Nataša Toplak and Tadej Avcin – Department of Allergology, Rheumatology and Clinical Immunology, University Children’s Hospital, University Medical Centre Ljubljana, Ljubljana, Slovenia) – pp 93-102 of “Vaccines and Autoimmunity” by Yehuda Shoenfeld et al. Wiley Ed. 2015.
Sovrastimolazione immunitaria. È in atto da anni un'”epidemia” di patologie infiammatorie che è sovrapponibile alla iatrogenicità causata dalla sovrastimolazione del sistema immunitario attraverso i vaccini. Ci sono ampie prove di collegamenti fra l’immunizzazione vaccinale e l’epidemia di diabete di tipo 1. Dei dati più recenti indicano che l’obesità, il diabete di tipo 2 e di altri componenti della sindrome metabolica sono fortemente associati con l’immunizzazione vaccinale e possono essere espressione diretta della risposta negativa del sistema immunitariocome reazione al sovraccarico immunitario.
Patologie Autoimmuni da Sovrastimolazione Vaccinale
L’“epidemia” di diabete/prediabete sembra accelerare nel momento in cui la prevalenza dell’obesità è stabilizzata, indicando che il sistema di risposta negativo del sistema immunitario è stato sopraffatto.
La teoria dell’induzione del sovraccarico immunitario causata dalle vaccinazioni può spiegare le osservazioni chiave che hanno confuso molte ipotesi tra loro in competizione.
Le ricerche indicate in questa rassegna evidenziano che il vaccino ha indotto un sovraccarico immunitario e spiega l’accentuarsi della divergenza tra l’aumento di prediabete e la steatosi epatica non alcolica in un momento in cui il epidemia di obesità è in calo nei bambini.
Fonte: Molecular and Genetic Medicine Classen JB, J Mol Genet Med 2014, S1:025
Review of Vaccine Induced Immune Overload and the Resulting Epidemics of Type 1 Diabetes and Metabolic Syndrome, Emphasis on Explaining the Recent Accelerations in the Risk of Prediabetes and other Immune Mediated Diseases. J Barthelow Classen MD* 3637 Rockdale Road, Manchester, MD 21102, USA *Corresponding author: J Barthelow Classen, 3637 Rockdale Road, Manchester, MD 21102, USA, Tel: 410-377-8526; E-mail: classen@vaccines.net
La teoria del sovraccarico immunitario indotto da vaccino spiega le epidemie parallele di diverse malattie autoimmuni. È noto che la patofisiologia è condivisa in molte malattie autoimmuni e infiammatorie.
I pazienti con malattia autoimmune spesso hanno più di una malattia autoimmune o hanno una storia familiare di più malattie autoimmuni. Non è quindi sorprendente che molte malattie infiammatorie aumentino insieme al diabete di tipo 1, infatti è previsto. È stata segnalata una crescente varietà di malattie nei bambini. Non ci sono dati sufficienti per sapere se la prevalenza della maggior parte delle malattie infiammatorie è in aumento.
Tuttavia, dato il numero e la varietà di malattie che sono state segnalate come in aumento nei bambini, è probabile che aumentino anche molte altre malattie.
Gli studi epidemiologici mostrano uno stretto legame tra il diabete di tipo 1 e altre malattie autoimmuni. Il diabete di tipo 1 è fortemente legato ad altre malattie autoimmuni nella sindrome autoimmune poliglandolare di tipo II [19]. In questa sindrome il 52% dei pazienti ha il diabete mellito, il 69% ha una malattia autoimmune della tiroide e il 100% ha la malattia di Addison.
I pazienti con diabete di tipo 1 e i loro parenti stretti sono a maggior rischio di malattia autoimmune organo-specifica [20].
Alcuni dati epidemiologici provengono da studi condotti su famiglie in cui diversi membri hanno una malattia autoimmune.
Gli studi familiari indicano che il diabete di tipo 1 è legato allo sviluppo di diverse malattie autoimmuni, tra cui malattie autoimmuni specifiche degli organi e malattie reumatoidi.
Parenti stretti di pazienti con diabete di tipo 1 hanno un rischio aumentato di un’ampia varietà di diversi autoanticorpi [21,22].
È stato scoperto che, a seconda della famiglia, il diabete di tipo 1 è associato a un aumento del rischio di una malattia autoimmune organo specifica o di una malattia reumatoide [23].
Un ampio studio sui mennoniti ha mostrato un legame tra il diabete di tipo 1 e altre malattie autoimmuni tra cui organo specifico e reumatoidi.
Uno studio italiano ha trovato che 14 su 11.241 pazienti trattati con interferone alfa hanno sviluppato diabete mellito [29]. L’interferone alfa aumenta anche il rischio di malattie autoimmuni specifiche per organo come la tiroidite e malattie reumatiche autoimmuni come il LES, l’artrite reumatoide, la psoriasi e il sarcoide [30].
È stato riportato che, in seguito alla somministrazione di interferone alfa, lo stesso paziente ha sviluppato entrambe le malattie autoimmuni reumatoide e organo specifica [31,32].
Bibliografia dell’aggiornamento del 1° Febbraio 2019
Al convegno internazionale sulle patologie autoimmuni, tenutosi a Lipsia nell’aprile 2016, la D.ssa Luisa Eca Guimarares tiene la 14ª sessione del Simposio sui Vaccini e spiega delle cose molto interessanti sui fisiologismi di induzione delle patologie autoimmuni da parte degli adiuvanti Alluminio presenti nei vaccini pediatrici e per adulti. Il video è scaricabile direttamente dalla pagina http://territorioscuola.com/videores/watch/mfraYYGYDA8 selezionando “Download” (se non riesci a scaricare, aggiorna la pagina e riprova).
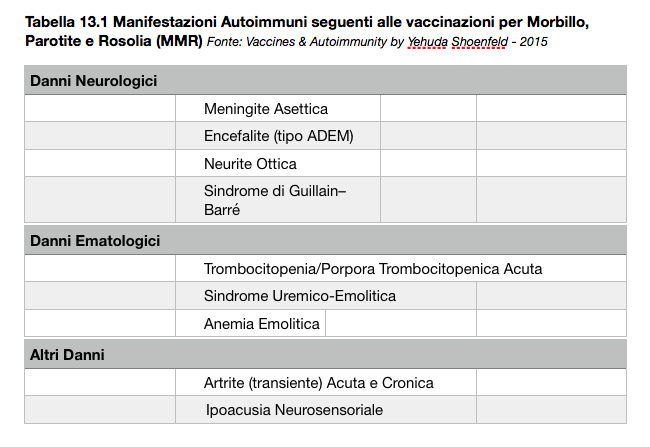
Tabella 13.1 Manifestazioni Autoimmuni seguenti alle vaccinazioni per Morbillo, Parotite e Rosolia (MMR)
“….La maggior parte degli eventi avversi che si manifestano dopo la vaccinazione MMR (Morbillo, Parotite e Rosolia) coinvolgono il sistema nervoso, le articolazioni e il sangue. Vedi Tabella 13.1. È da notare che tali tipi di patologia riflettono lo spettro delle manifestazioni autoimmuni che possono verificarsi a seguito delle infezioni virali da MMR (Agmon-Levin et al., 2009b). Esiste un certo numero di meccanismi plausibili che potrebbe spiegare l’autoimmunità indotta dal vaccino, compreso il fatto che i vaccini MMR sono composti da antigeni infettivi, adiuvanti immunitari, conservanti e altri ingredienti che possono innescare lo sviluppo o l’esacerbazione di fenomeni autoimmuni (Colafrancesco et al.,2013). I vaccini sono una parte essenziale della prevenzione della medicina moderna, ma il loro potenziale di innescare la sindrome ASIA in soggetti sensibili non dovrebbe essere ignorato. Ulteriori sforzi dovrebbero anche essere fatti per indagare più a lungo termine la sicurezza dei vaccini comunemente impiegati (Shoenfeld e Agmon-Levin, 2011; Perricone et al., 2013)….”
“….L’analisi Istopatologica ha confermato l’encefalite; l’analisi immunoistochimica e il qPCR sono risultati positivi per il virus della rosolia su tessuto cerebrale. Il virus è stato isolato anche dal liquido cerebrospinale (CSF) e da altri campioni clinici. La sequenza ottenuta dal virus isolato era identica a quella del ceppo vaccinale RA 27/3, suggerendo un legame causale (Gualberto et al., 2013)….”
Patologie Autoimmuni Causate dal Vaccino MMR
“…There are several lines of evidence that thrombocytopenia is related to the MMR vaccination (Molina and Shoenfeld, 2005). In the US Vaccine Adverse Event Reporting System (VAERS), there were 250 reports (259 events) of thrombocytopenia within 77 days post-vaccination (Mantadakis et al., 2010). Usually, the onset appears to be sudden, with a median of 13 days. Patient age was largely variable, ranging from 16 weeks to 53 years…”
“…Ci sono diverse prove che la trombocitopenia è legata alla vaccinazione MMR (Molina e Shoenfeld, 2005). Negli Stati Uniti il Vaccine Adverse Event Reporting System (VAERS), segnala 250 rapporti (259 eventi) di trombocitopenia entro 77 giorni dopo la vaccinazione (Mantadakis et al., 2010). Solitamente, l’insorgenza sembra essere improvvisa, con una media di 13 giorni. L’età del paziente era in gran parte variabile, da 16 settimane a 53 anni…”
MMR – Una Triade per la Malattia Autoimmune
Fonte: “Vaccines & Autoimmunity” di Yehuda Shoenfeld, Nancy Agmon-Levin, Lucija Tomljenovic. Edizione in lingua inglese – 2015 – Editore Wiley Blackwell.
Alla luce dei numerosi lavori pubblicati dal Dott. Yehuda Shoenfeld nel Dicembre 2015 nel Testo Unico “Vaccines & Autoimmunity”, è confermatissima la responsabilità di numerosi vaccini per uso pediatrico e adulto nello scatenare, anche dopo anni dall’inoculo, numerosi tipi di reazioni autoimmuni in cui l’organismo produce degli auto-anticorpi in grado di attaccare “senza fine” sé stesso… L’abstract in immagine, peraltro pubblicato da ELSEVIER, maggior editore medico mondiale, si riferisce ad un lavoro precedente di Soriano, Nesher e dello stesso Shoenfeld uscito nel 2014 e di cui è possibile trovare gli originali nei links sotto riportati. Già prima del 2014, tuttavia, erano state riportate numerose osservazioni da parte di altri autori su questa strettissima correlazione. Assurdo il fatto che i periti superpartes, come coloro che si occupano di giustizia nelle cause per danni da vaccino non siano adusi a consultare queste pietre miliari della ricerca sull’autoimmunologia. Intanto ci sono ancora moltissimi genitori di bambini danneggiati da vaccinazioni “a tappeto” che non sospettano minimamente la causa vaccinale come scatenante la malattia autoimmune subìta dai loro bambini…
Ci sarà pure una ragione del fatto che nel testo (altamente referenziato e peer-reviewed) “Vaccines & Autoimmunity” di Shoenfeld, Tomljenovic, Agmon-Levin – 2015, a chiusura del capitolo introduttivo (pagina 6), viene riportato a chiare lettere che le varie tipologie di vaccini attualmente in commercio possono scatenare delle reazioni autoimmuni a breve-medio e lungo termine?
Vaccine can induce Autoimmune Reactions
Gli autori (non sono solo loro tre ma diverse decine hanno contribuito alla stesura di questo libro di testo sulle patologie autoimmuni), pur sostenendo la validità delle vaccinazioni, avvertono i medici e i clinici di tutto il mondo che il rischio (anche mortale) esiste. Dato ciò, come mai i pediatri e gli altri medici italiani IGNORANO COMPLETAMENTE i risultati di questi studi clinici ultratrentennali? Come mai continuano a vaccinare i neonati, i bambini, senza nemmeno considerare le loro specificità?