
Stephanie Seneff, Greg Nigh, Anthony M. Kyriakopoulos, Peter A. McCullough
Traduzione e adattamento a cura di Davide Suraci
Introduzione
In questo documento, richiamiamo l’attenzione su tre aspetti molto importanti del profilo di sicurezza di queste vaccinazioni. Il primo è la soppressione, ampiamente documentata, dell’immunità innata principalmente attraverso la soppressione dell’IFN-α e la sua cascata di segnalazione associata. Questa soppressione avrà una vasta gamma di conseguenze tra cui, non ultima, la riattivazione di infezioni virali latenti e la ridotta capacità di combattere efficacemente future infezioni. Il secondo è la disregolazione del sistema sia per prevenire che per rilevare la trasformazione maligna geneticamente guidata all’interno delle cellule e il conseguente potenziale per la vaccinazione di promuovere tali trasformazioni. In terzo luogo, la vaccinazione con mRNA interrompe potenzialmente la comunicazione intracellulare effettuata dagli esosomi e induce le cellule che assorbono l’mRNA della glicoproteina spike a produrre alti livelli di esosomi che trasportano la glicoproteina spike, con conseguenze infiammatorie potenzialmente gravi.
Generalità
Le numerose alterazioni dell’mRNA del vaccino nascondono l’mRNA dalle difese cellulari e promuovono un’emivita biologica più lunga e un’elevata produzione di proteina spike. Tuttavia, la risposta immunitaria al vaccino è molto diversa da quella a una vera una vera infezione da SARS-CoV-2. In questo documento, presentiamo le prove che la vaccinazione induce una profonda compromissione della segnalazione dell’interferone di tipo I, che ha diverse conseguenze negative per la salute umana. Le cellule immunitarie che hanno assorbito le nanoparticelle del vaccino rilasciano in circolazione un gran numero di esosomi contenenti proteine spike insieme a microRNA (miRNA) critici che inducono una risposta di segnalazione nelle cellule riceventi in siti distanti. Identifichiamo anche potenziali profondi disturbi nel controllo normativo della sintesi proteica e dei sistemi di controllo del cancro. Questi disturbi hanno potenzialmente un nesso causale con la malattia neurodegenerativa, la miocardite, la trombocitopenia immunitaria, la paralisi di Bell, le malattie del fegato, l’immunità adattativa compromessa, la risposta al danno del DNA compromessa e la tumorigenesi. Mostriamo prove dal database VAERS a sostegno della nostra ipotesi. Riteniamo che una valutazione completa del rapporto rischio/beneficio dei vaccini a mRNA li metta in discussione come contributori positivi alla salute pubblica.

Questo paper riassume l’attuale letteratura sull’mRNA e sui suoi effetti sulla biologia molecolare all’interno delle cellule umane. Riconosciamo che esiste un’ampia gamma di opinioni in questa fase nascente della tecnologia dell’mRNA. Dato il suo diffuso dispiegamento prima del lavoro di base su tanti dei meccanismi di cui discutiamo qui, riteniamo che il nostro lavoro sia importante per fornire un’ampia comprensione delle revisioni presenti e future relative al fiorente lavoro molecolare preclinico svolto in quest’area.
In questo articolo, viene analizzata la letteratura scientifica che suggerisce che la vaccinazione con un vaccino a mRNA avvia una serie di eventi biologici che non solo sono diversi da quelli indotti dall’infezione, ma sono in molti modi dimostrabilmente controproducenti per la competenza immunitaria sia a breve che a lungo termine e normale funzione cellulare. Queste vaccinazioni hanno ora dimostrato di sottoregolare i percorsi critici relativi alla sorveglianza del cancro, al controllo delle infezioni e all’omeostasi cellulare. Introducono nel corpo materiale genetico altamente modificato. Una prestampa ha rivelato una notevole differenza tra le caratteristiche della risposta immunitaria a un’infezione da SARS-CoV-2 rispetto alla risposta immunitaria a un vaccino mRNA contro COVID-19 (Ivanova et al., 2021).
L’analisi dell’espressione genica differenziale delle cellule dendritiche periferiche ha rivelato una drammatica sovraregolazione degli interferoni di tipo I e II (IFN) nei pazienti con COVID-19, ma non nei vaccinati.
Un’osservazione notevole che hanno fatto è stata che c’era un’espansione delle cellule staminali e progenitrici ematopoietiche circolanti (HSPC) nei pazienti COVID-19, ma questa espansione era notevolmente assente dopo la vaccinazione. Anche una sorprendente espansione dei plasmablasti circolanti osservata nei pazienti COVID-19 non è stata osservata nei vaccinati. Tutte queste osservazioni sono coerenti con l’idea che i vaccini anti-COVID-19 sopprimono attivamente la segnalazione IFN di tipo I, come discuteremo di seguito. In questo documento ci concentreremo ampiamente, anche se non esclusivamente, sulla soppressione dell’IFN di tipo I indotta dalla vaccinazione e sulla miriade di effetti a valle che ciò ha sulla relativa cascata di segnalazione.
I vaccini a mRNA prodotti da Pfizer/BioNTech e Moderna sono stati visti come un aspetto essenziale degli sforzi per controllare la diffusione di COVID-19…
I governi sono reticenti a considerare la possibilità che queste “vaccinazioni” a mRNA possano causare danni in modi inaspettati, e soprattutto che tali danni possano persino superare i benefici ottenuti nella protezione da malattie gravi.
È ora chiaro che gli anticorpi indotti dai vaccini svaniscono in appena 3-10 settimane dopo la seconda dose (Shrotri et al., 2021), tanto che alle persone viene consigliato di farsi iniezioni di richiamo a intervalli regolari (Centers for Disease Controllo e prevenzione, 2021b). È anche diventato evidente che varianti in rapida evoluzione come il ceppo Delta e ora Omicron stanno mostrando resistenza agli anticorpi indotti dai vaccini, attraverso mutazioni nella proteina spike (Yahi et al., 2021). Inoltre, è diventato chiaro che i vaccini non prevengono la trasmissione della malattia, ma si può solo affermare che riducano la gravità dei sintomi (Kampf, 2021a). Uno studio che ha confrontato i tassi di vaccinazione con i tassi di infezione da COVID-19 in 68 paesi e 2947 contee negli Stati Uniti all’inizio di settembre 2021, non ha trovato alcuna correlazione tra i due, suggerendo che questi vaccini non proteggono dalla diffusione della malattia (Subramanian e Kumar, 2947). Per quanto riguarda la gravità dei sintomi, anche questo aspetto comincia a essere messo in dubbio, come dimostra un focolaio in un ospedale israeliano che ha portato alla morte di cinque pazienti ospedalieri completamente vaccinati (Shitrit et al., 2021). Allo stesso modo, Brosh-Nissimov et al. (2021) hanno riferito che 34/152 (22%) dei pazienti completamente vaccinati tra 17 ospedali israeliani sono morti di COVID-19.
La crescente evidenza che i vaccini fanno poco per controllare la diffusione della malattia e che la loro efficacia diminuisce nel tempo rende ancora più imperativo valutare il grado in cui i vaccini potrebbero causare danni. Che le vaccinazioni con mRNA della proteina spike modificata da SARS-CoV-2 abbiano impatti biologici è fuori discussione. Qui tentiamo di distinguere quegli impatti da vaccini a mRNA da quelli derivanti dall’infezione naturale e di stabilire un quadro meccanicistico che colleghi quegli impatti biologici unici alle patologie ora associate alla vaccinazione. Riconosciamo che i collegamenti causali tra gli effetti biologici avviati dalla vaccinazione mRNA e gli esiti avversi non sono stati stabiliti nella grande maggioranza dei casi.
Il Ruolo degli IFN di Tipo 1
Gli IFN di tipo I svolgono un ruolo importante nella risposta immunitaria a molteplici fattori di stress. Infatti, hanno goduto di un valore terapeutico clinico come opzione terapeutica per una varietà di malattie e condizioni, tra cui infezioni virali, tumori solidi, disturbi mieloproliferativi, neoplasie ematopoietiche e malattie autoimmuni come la sclerosi multipla (Passegu e Ernst, 2009).
Come gruppo, gli IFN svolgono ruoli estremamente complicati e pleiotropici che sono coordinati e regolati attraverso l’attività della famiglia dei fattori regolatori IFN, o IRF (Kaur e Fang, 2020). IRF9 è più direttamente coinvolto nell’immunità antivirale e antitumorale e nella regolazione genetica (Alsamman e El-Masry, 2018; Huang et al., 2019; Zitvogel et al., 2015).
Strettamente correlate a questo sono le cellule dendritiche plasmacitoidi (pDC), un raro tipo di cellula immunitaria che circola nel sangue ma migra verso gli organi linfoidi periferici durante un’infezione virale. Rispondono a un’infezione virale aumentando nettamente la produzione di IFN di tipo I. L’IFN-α rilasciato nei linfonodi induce le cellule B a differenziarsi in plasmablasti. Successivamente, l’interleuchina-6 (Il-6) induce i plasmablasti ad evolversi in plasmacellule che secernono anticorpi (Jego et al., 2003). Pertanto, gli IFN svolgono un ruolo fondamentale sia nel controllo della proliferazione virale sia nell’induzione della produzione di anticorpi. Centrale sia per l’immunità antivirale che per quella antitumorale, l’IFN-α è prodotto da macrofagi e linfociti quando viene colpito da infezioni virali o batteriche o incontra cellule tumorali (De Andrea et al., 2002). Il suo ruolo come potente terapia antivirale è stato riconosciuto nel trattamento delle complicanze del virus dell’epatite C (Feng et al., 2012), dell’infezione da citomegalovirus (Delannoy et al., 1999), dell’infezione cronica da virus ebola attivo (Sakai et al., 1998 ), malattia infiammatoria intestinale associata a infezione da herpes virus (Ruther et al., 1998) e altri.
Alterazione del Signaling Antitumorale dell’IFN-1
La segnalazione alterata dell’IFN di tipo I è collegata a molti rischi di malattia, in particolare il cancro, poiché la segnalazione dell’IFN di tipo I sopprime la proliferazione sia dei virus che delle cellule tumorali arrestando il ciclo cellulare, in parte attraverso la sovraregolazione di p53, un gene oncosoppressore e varie cicline -inibitori della chinasi dipendenti (Musella et al., 2017; Matsuoka et al., 1998). L’IFN-α induce anche la presentazione dell’antigene di classe 1 di maggiore istocompatibilità (MHC) da parte delle cellule tumorali, facendole riconoscere più facilmente dal sistema di sorveglianza del cancro (Heise et al., 2016; Sundstedt et al., 2008). La gamma di effetti antitumorali avviati dall’espressione di IFN-α è sbalorditiva e si verifica attraverso meccanismi sia diretti che indiretti. Gli effetti diretti includono l’arresto del ciclo cellulare, l’induzione della differenziazione cellulare, l’inizio dell’apoptosi, l’attivazione di natural killer e cellule T CD8+ e altri (Schneider et al., 2014).
“Vaccini” a mRNA Non Conformi al Controllo di Qualità Cellulare
Questo studio (McKernan et al., 2021) ha dimostrato che esiste un significativo arricchimento del contenuto di GC (Guanina e Citosina) negli mRNA dei vaccini (53% in BNT162b2 e 61% in Moderna mRNA-1273) rispetto all’mRNA nativo di SARS-CoV-2 (36%). Il contenuto arricchito di GC degli mRNA è il risultato dell’ottimizzazione del codone eseguita durante lo sviluppo degli mRNA utilizzati nei vaccini SARS-CoV-2, apparentemente senza determinare l’effetto sulle strutture secondarie, in particolare la formazione del quadruplex della guanina (G quadruplex).
Non Ottimizzazione dei Codoni e Produzione di Proteine Aliene
L’ottimizzazione del codone descrive la produzione di polipeptidi e proteine sintetici ottimizzati per il codone utilizzati nelle terapie biotecnologiche (come gli mRNA sintetici utilizzati per la vaccinazione SARS-CoV-2). Le assegnazioni alterate del codone all’interno del modello di mRNA aumentano notevolmente la quantità di polipeptidi e/o proteine prodotte (Mauro e Chappell, 2014). La sostituzione del codone sinonimo comporta anche un cambiamento nei ruoli normativi e strutturali multifunzionali delle proteine risultanti (Shabalina et al., 2013). Per questo motivo, l’ottimizzazione del codone è stata messa in guardia a causa dei suoi conseguenti cambiamenti che causano perturbazioni nella conformazione secondaria dei prodotti proteici con effetti potenzialmente devastanti sulla loro risultante immunogenicità, efficacia e funzione (Zhou et al., 2013; Agashe et al., 2013 ). In particolare, varie malattie umane sono il risultato di polimorfismi nucleotidici sinonimi (McCarthy et al., 2017).
Implicazioni Inquietanti – Compromissione dei Blocchi Protettivi Anticancro
La glicoproteina spike SARS-CoV-2 modifica la produzione di esosomi della cellula ospite. La trasfezione delle cellule con il gene della proteina spike e la successiva produzione della proteina spike SARS-CoV-2 si traduce in quelle cellule che generano esosomi contenenti microRNA che sopprimono la produzione di IRF9 mentre attivano una serie di trascrizioni geniche pro-infiammatorie (Mishra e Banerjea, 2021). Poiché questi vaccini sono specificamente progettati per indurre una produzione elevata e continua di glicoproteine spike SARS-CoV-2, le implicazioni sono inquietanti. Come descritto sopra, l’inibizione di IRF9 sopprimerà TRAIL(Trans-Allegheny Interstate Line) e tutti i suoi effetti regolatori e a valle che inducono l’apoptosi. Ci si dovrebbe anche aspettare che la soppressione dell’IRF9 tramite microRNA esosomiale comprometta gli effetti protettivi contro il cancro dell’attività del gene BRCA2, che dipende da quella molecola per la sua attività come descritto sopra. I tumori associati a BRCA2 includono cancro al seno, alle tube di Falloppio e alle ovaie per le donne, cancro alla prostata e al seno per gli uomini, leucemia mieloide acuta nei bambini e altri (National Cancer Institute, 2021).
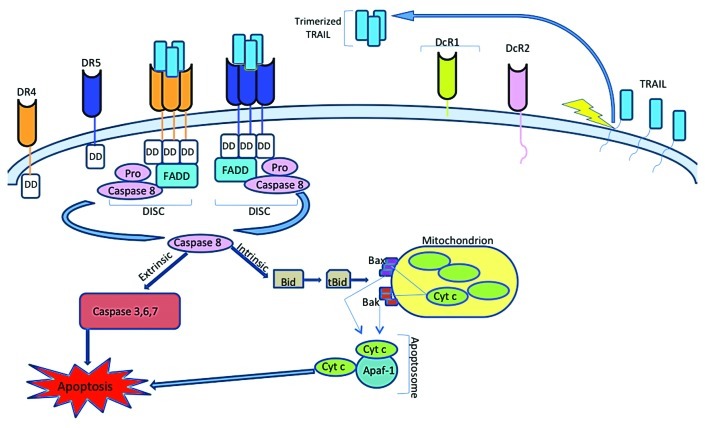
È stato anche dimostrato che la vaccinazione sopprime sia IRF7 che STAT2 (Liu et al., 2021). Ci si può aspettare che ciò interferisca con gli effetti protettivi contro il cancro di BRCA1 come descritto sopra. I tumori associati all’attività BRCA1 compromessa comprendono il cancro al seno, all’utero e alle ovaie nelle donne; cancro alla prostata e al seno negli uomini; e un modesto aumento del cancro al pancreas sia per gli uomini che per le donne (rischio di cancro e gene BRCA1, 2021).
Riattivazione della Varicella-Zoster
La segnalazione del recettore IFN di tipo I nelle cellule T CD8+ è fondamentale per la generazione di cellule effettrici e di memoria in risposta a un’infezione virale (Kolumam et al., 2005). Le cellule T CD8+ possono bloccare la riattivazione dell’infezione latente da herpes nei neuroni sensoriali (Liu et al., 2000). Se la segnalazione dell’IFN di tipo I è compromessa, come accade dopo la vaccinazione ma non dopo l’infezione naturale da SARS-CoV-2, anche la capacità delle cellule T CD8+ di tenere sotto controllo l’herpes sarebbe compromessa. Potrebbe essere questo il meccanismo al lavoro in risposta ai vaccini?
Trombocitopenia Immunitaria
La trombocitopenia immunitaria è una malattia autoimmune, in cui il sistema immunitario attacca le piastrine circolanti. La porpora trombocitopenica immunitaria (ITP) è stata associata a diverse vaccinazioni, tra cui morbillo, parotite, rosolia (MMR), epatite A, varicella, difterite, tetano, pertosse (DPT), poliomielite orale e influenza (Perricone et al., 2014). Sebbene vi sia un’ampia consapevolezza che i vaccini basati sul DNA dell’adenovirus possono causare trombocitopenia trombotica immunitaria indotta da vaccino (VITT) (Kelton et al., 2021), i vaccini a mRNA non sono privi di rischi per la VITT, poiché sono stati pubblicati casi di studio che documentano tale eventi, inclusa la trombosi del seno venoso cerebrale mortale e fatale (Lee et al., 2021; Akiyama et al., 2021; Atoui et al., 2022; Zakaria et al., 2021). Si ritiene che il meccanismo coinvolga gli anticorpi VITT che si legano al fattore piastrinico 4 (PF4) e formino immunocomplessi che inducono l’attivazione piastrinica. Le successive cascate di coagulazione causano la formazione di microcoaguli diffusi nel cervello, nei polmoni, nel fegato, nelle gambe e altrove, associati a un drastico calo della conta piastrinica (Kelton et al., 2021). La reazione al vaccino è stata descritta come molto simile alla trombocitopenia indotta da eparina (HIT), tranne per il fatto che la somministrazione di eparina non è coinvolta (Cines e Bussel, 2021). (Fine prima parte)
Raccomando la diffusione capillare della presente sintesi che potrete trovare in versione integrale direttamente da qui: “Innate immune suppression by SARS-CoV-2 mRNA vaccinations: The role of G-quadruplexes, exosomes, and MicroRNAs”





